Download
1 / 1
10 likes | 132 Views
Physical Mapping of Compactum and Three Pistil Loci on Wheat Chromosome 2D Using Radiation Hybrids. DBI-0822100. Vijay K. Tiwari 1 , Kasandra Lopez 1 , Oscar Riera-Lizarazu 2 , Shahryar F. Kianian 3 , Jeffrey M. Leonard 1.

E N D
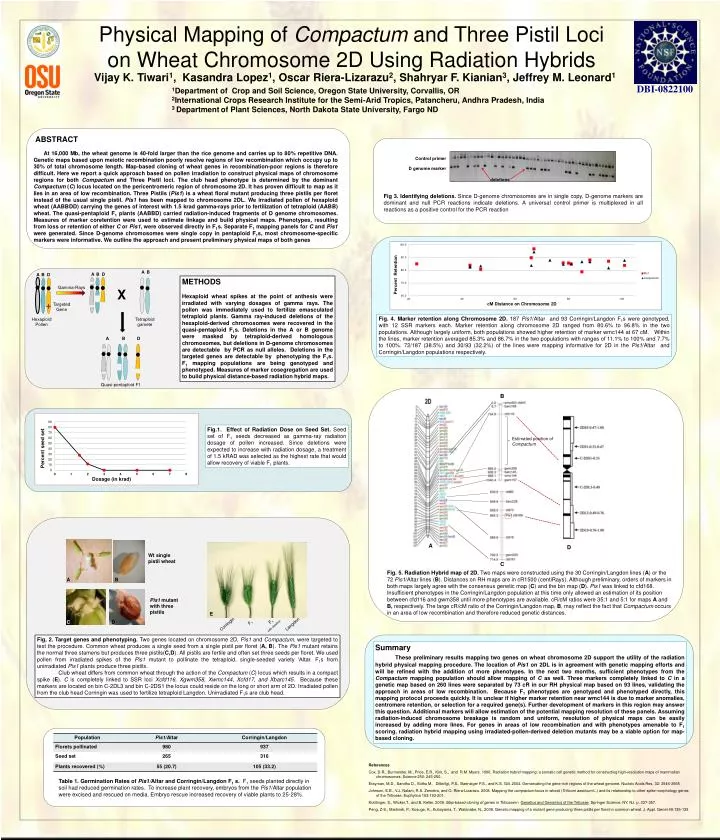
Physical Mapping of Compactum and Three Pistil Loci on Wheat Chromosome 2D Using Radiation Hybrids DBI-0822100 Vijay K. Tiwari1, Kasandra Lopez1, Oscar Riera-Lizarazu2, Shahryar F. Kianian3, Jeffrey M. Leonard1 1Department of Crop and Soil Science, Oregon State University, Corvallis, OR 2International Crops Research Institute for the Semi-Arid Tropics, Patancheru, Andhra Pradesh, India 3 Departmentof Plant Sciences, North Dakota State University, Fargo ND ABSTRACT At 16,000 Mb, the wheat genome is 40-fold larger than the rice genome and carries up to 80% repetitive DNA. Genetic maps based upon meiotic recombination poorly resolve regions of low recombination which occupy up to 30% of total chromosome length. Map-based cloning of wheat genes in recombination-poor regions is therefore difficult. Here we report a quick approach based on pollen irradiation to construct physical maps of chromosome regions for both Compactum and Three Pistil loci. The club head phenotype is determined by the dominant Compactum (C) locus located on the pericentromeric region of chromosome 2D. It has proven difficult to map as it lies in an area of low recombination. Three Pistils (Pis1) is a wheat floral mutant producing three pistils per floret instead of the usual single pistil. Pis1 has been mapped to chromosome 2DL. We irradiated pollen of hexaploid wheat (AABBDD) carrying the genes of interest with 1.5 krad gamma-rays prior to fertilization of tetraploid (AABB) wheat. The quasi-pentaploid F1 plants (AABBD) carried radiation-induced fragments of D genome chromosomes. Measures of marker coretention were used to estimate linkage and build physical maps. Phenotypes, resulting from loss or retention of either C or Pis1, were observed directly in F1s. Separate F1 mapping panels for C and Pis1 were generated. Since D-genome chromosomes were single copy in pentaploid F1s, most chromosome-specific markers were informative. We outline the approach and present preliminary physical maps of both genes deletions METHODS Hexaploid wheat spikes at the point of anthesis were irradiated with varying dosages of gamma rays. The pollen was immediately used to fertilize emasculated tetraploid plants. Gamma ray-induced deletions of the hexaploid-derived chromosomes were recovered in the quasi-pentaploid F1s. Deletions in the A or B genome were masked by tetraploid-derived homologous chromosomes, but deletions in D-genome chromosomes are detectable by PCR as null alleles. Deletions in the targeted genes are detectable by phenotyping the F1s. F1 mapping populations are being genotyped and phenotyped. Measures of marker cosegregation are used to build physical distance-based radiation hybrid maps. Gamma-Rays X A B A B D Targeted Gene A B D Fig. 4. Marker retention along Chromosome 2D. 187Pis1/Altar and 93 Corringin/Langdon F1s were genotyped, with 12 SSR markers each. Marker retention along chromosome 2D ranged from 80.6% to 96.8% in the two populations. Although largely uniform, both populations showed higher retention of marker wmc144 at 67 cM. Within the lines, marker retention averaged 85.3% and 86.7%in the two populations with ranges of 11.1% to 100% and 7.7% to 100%. 72/187 (38.5%) and 30/93 (32.2%) of the lines were mapping informative for 2D in the Pis1/Altar and Corringin/Langdon populations respectively. Hexaploid Pollen Tetraploid gamete A B D Quasi-pentaploid F1 B A Fig.1. Effect of Radiation Dose on Seed Set. Seed set of F1 seeds decreased as gamma-ray radiation dosage of pollen increased. Since deletions were expected to increase with radiation dosage, a treatment of 1.5 kRAD was selected as the highest rate that would allow recovery of viable F1 plants. Estimated position of Compactum A D C C D B Fig. 5. Radiation Hybrid map of 2D. Two maps were constructed using the 30 Corringin/Langdon lines (A) or the 72 Pis1/Altar lines (B). Distances on RH maps are in cR1500 (centiRays). Although preliminary, orders of markers in both maps largely agree with the consensus genetic map (C) and the bin map (D). Pis1 was linked to cfd168. Insufficient phenotypes in the Corringin/Langdon population at this time only allowed an estimation of its position between cfd116 and gwm358 until more phenotypes are available. cR/cM ratios were 35:1 and 5:1 for maps A and B, respectively. The large cR/cM ratio of the Corringin/Langdon map, B, may reflect the fact that Compactum occurs in an area of low recombination and therefore reduced genetic distances. Wt single pistil wheat Summary These preliminary results mapping two genes on wheat chromosome 2D support the utility of the radiation hybrid physical mapping procedure. The location of Pis1 on 2DL is in agreement with genetic mapping efforts and will be refined with the addition of more phenotypes. In the next two months, sufficient phenotypes from the Compactum mapping population should allow mapping of C as well. Three markers completely linked to C in a genetic map based on 260 lines were separated by 73 cR in our RH physical map based on 93 lines, validating the approach in areas of low recombination. Because F1 phenotypes are genotyped and phenotyped directly, this mapping protocol proceeds quickly. It is unclear if higher marker retention near wmc144 is due to marker anomalies, centromere retention, or selection for a required gene(s). Further development of markers in this region may answer this question. Additional markers will allow estimation of the potential mapping resolution of these panels. Assuming radiation-induced chromosome breakage is random and uniform, resolution of physical maps can be easily increased by adding more lines. For genes in areas of low recombination and with phenotypes amenable to F1 scoring, radiation hybrid mapping using irradiated-pollen-derived deletion mutants may be a viable option for map-based cloning. A B Table 1. Germination Rates of Pis1/Altar and Corringin/Langdon F1 s. F1 seeds planted directly in soil had reduced germination rates. To increase plant recovery, embryos from the Pis1/Altar population were excised and rescued on media. Embryo rescue increased recovery of viable plants to 25-28%. Pis1 mutant with three pistils E Fig 3. Identifying deletions. Since D-genome chromosomes are in single copy, D-genome markers are dominant and null PCR reactions indicate deletions. A universal control primer is multiplexed in all reactions as a positive control for the PCR reaction F1 F1 with deletion Corringin Langdon C D Fig, 2. Target genes and phenotyping. Two genes located on chromosome 2D, Pis1 and Compactum, were targeted to test the procedure.Common wheat produces a single seed from a single pistil per floret (A, B). The Pis1 mutant retains the normal three stamens but produces three pistils(C,D). All pistils are fertile and often set three seeds per floret. We used pollen from irradiated spikes of the Pis1 mutant to pollinate the tetraploid, single-seeded variety ‘Altar. F1s from unirradiatedPis1 plants produce three pistils. Club wheat differs from common wheat through the action of the Compactum (C) locus which results in a compact spike (E). C is completely linked to SSR loci Xcfd116, Xgwm358, Xwmc144, Xcfd17, and Xbarc145. Because these markers are located on bin C-2DL3 and bin C-2DS1 the locus could reside on the long or short arm of 2D. Irradiated pollen from the club head Corringin was used to fertilize tetraploid Langdon. Unirradiated F1s are club head. Control primer D genome marker References Cox, D.R., Burmeister, M., Price, E.R., Kim, S., and R.M. Myers. 1990. Radiation hybrid mapping: a somatic cell genetic method for constructing high-resolution maps of mammalian chromosomes. Science 250: 245-250. Erayman, M.D., Sandhu D., Sidhu M., Dilbirligi, P.S., Baenziger P.S., and K.S. Gill. 2004. Demarcating the gene-rich regions of the wheat genome. Nucleic Acids Res. 32: 3546-3565 Johnson, E.B., V.J. Nalam, R.S. Zemetra, and O. Riera-Lizarazu. 2008. Mapping the compactum locus in wheat (Triticumaestivum L.) and its relationship to other spike morphology genes of the Triticeae. Euphytica 163:193-201. Krattinger, S., Wicker,T., and B. Keller. 2009. Map-based cloning of genes in Triticeaein Genetics and Genomics of the Triticeae. Springer Science, NY, NJ. p. 337-357. Peng, Z-S., Martinek, P., Kosuge, K., Kuboyama, T., Watanabe, N., 2008. Genetic mapping of a mutant gene producing three pistils per floret in common wheat. J. Appl. Genet 49:135-139