Download
1 / 40
450 likes | 767 Views
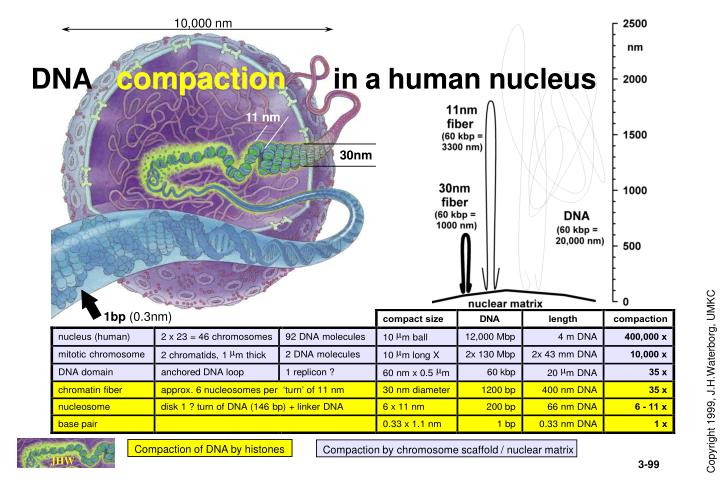
Compaction by chromosome scaffold / nuclear matrix. 10,000 nm. DNA compaction in a human nucleus. 11 nm. 30nm. 1bp (0.3nm). Compaction of DNA by histones. H1. HISTONES are highly conserved, small, basic proteins. Linker histone. H2A. H2B. helix. Histone acetylation

E N D
Compaction by chromosome scaffold / nuclear matrix 10,000 nm DNA compactioninahuman nucleus 11 nm 30nm 1bp (0.3nm) Compaction of DNA by histones JHW
H1 HISTONESare highly conserved,small, basic proteins Linker histone H2A H2B helix • Histone acetylation • is a reversible modification • of lysines in the N-termini • of the core histones. • Result: • reduced binding to DNA • destabilization of chromatin Core histones variable H3 H4 conserved N JHW
Core Histones The histone-fold • The basic structure of ALL • core histones is the same: • 1 long hydrophobic alpha-helix, • bordered by • 2 short hydrophobic alpha helices • that form pairs • H2A - H2B and H3 - H4 • which interact. References:Moudrianakis et al. PNAS 88, 10138 (1991); PNAS 90, 10489 (1993); PNAS 92, 11170 (1995) JHW
Histone octamer assembly Histone octamer H3-H4 tetramer H2A-H2B dimer JHW
H4 H3 H2B H2A Nucleosome features • 146-149 bp DNA in a 1.65 turns of a flat, left-handed superhelix • one pseudo twofold axis centered at the “dyad” (reference: 0 helical turns) • one base-pair precisely at the dyad • sharp bends at + 1.5 and + 4-5 turns • Histone-fold domains organize 121 bp of DNA. The DNA is bound at 10 bp intervals through many contacts, including penetration of arginines at all 14 minor grooves facing the protein core • The grooves from neighboring DNA turns line up; forming channels • H3 and H2B N-termini exit one of these channels every 20bp. • The H4 tail establishes contacts with the next core particle. JHW
1 mM 5 mM Zhou, Gerchman, Ramakrishnan, Travers, Muyldermans Nature 395, 402 (1998) An, Leuba, van Holde, Zlatanova PNAS 95, 3396 (1998) Chromatosome C Core histone octamer + 1 Linker Histone + 2 full turns of DNA (168 bp) N H1 H3 H3 Linker Histone and histone termini control linker DNA entry/exit of chromatosome in chromatin fiber. JHW
Chromatinfibers 11 nm (beads) 30 nm chromatin fiber highly acetylated core histones (especially H3 and H4) + charged N termini (bind DNA on neigboring nucleosomes) • HIGH level of histone H1 • Reduced level of histone H1 • Gene transcription possible • NO gene transcription JHW
Alternative chromatin fiber models Zigzag fiber
Les bases moléculaires de l’épigénétique Altérations de la chromatine via des modifications d’histones et l’action des protéines chromatiniennes (HP1, Polycomb/Trithorax) Variants d’histones Méthylation de l’ADN Liens entre méthylation de l’ADN et modifications d’histones
Histone modifications 27 Cell (2002) 111, 285-91
5 16 20 8 12 Ac or Me Ac Ac Ac Ac Ac-S-G-R-G-K-G-G-K-G-L-G-K-G-G-A-K-R-H-R-K-V-L-R-D- + + + + + + + + + + 27 4 18 9 23 14 Ac or Me Me Ac Ac Ac Ac A-R-T-K-Q-T-A-R-K-S-T-G-G-K-A-P-R-K-Q-L-A-T-K-A-A-R-K-S-A-P- + + + + + + + + + - N - O O - - C C C g e - N+ P P - a d b C C C - O O O - - - - - - - N P - - C C N C C e - - - C C C C - O - - O - Acetylation of conserved lysines The N-termini of histones H4 and H3, and their acetylation patterns, are absolutely conserved. H4 N-terminus H3 N-terminus DNA backbone binding Lysine Acetyl-CoA HAT (Histone Acetyl-Transferase) Histone Deacetylase reversible reactions CoA e-N-Acetyl-Lysine no DNA binding JHW
Acétylation vs déacetylation des histones Chromatine décondensée: Etat transcriptionnel actif Acétylation des histones Chromatine condensée: Etat transcriptionnel réprimé Déacetylation des histones
Histone Methylation / Demethylation C C C C C C Histones can be methylated at lysines or arginines. Example: H3 K4 methylation N Lysine C C C g e N+ a d b C C C O S-adenosylmethyionine HMT (Histone Methyl-Transferase) Histone demethylase DNA backbone binding may not be strongly affected, but specific proteins may recognize these modifications N e-N-monomethyl-Lysine N+ C C C e C C C O S-adenosylmethyionine HMT (Histone Methyl-Transferase) Histone demethylase N Histone demethylases found for K4, K9, K27 and K36 e-N-dimethyl-Lysine N+ C C C e e S-adenosylmethyionine C C C O HMT (Histone Methyl-Transferase) Histone demethylase N N+ C C C e-N-trimethyl-Lysine e e C C C O
Méthylation vs déméthylation des histones Méthylation des lysines H3K4, H3K36 et H3K79 Chromatine décondensée: Etat transcriptionnel actif Déméthylation des lysines H3K9, H3K27 et H4K20 Méthylation des lysines H3K9, H3K27 et H4K20 Chromatine condensée: Etat transcriptionnel réprimé Déméthylation des lysines H3K4, H3K36 et H3K79
Les modifications d’histones peuvent être reconnues par d’autres protéines chromatiniennes : Le concept de « writer » et « reader »
HP1 Polycomb PC PRC1 Constitutive Heterochromatin Polycomb silencing (Facultative Heterochromatin) H3-K9me and H3-K27me recognition by the chromodomain proteins HP1 and Polycomb SUV39H1/2 (HMT) EZH1/2 (HMT) H3-K9me2/3 H3-K27me3
NURF-301 ISWI-dependant Chromatin remodeling leading to Chromatin opening and activation NURF 301 ISWI H3-K4me recognition by the PHD domain protein NURF-301 MLL1/2/3 TRX/ASH1 (HMT) H3-K4me3 Trithorax activation Wysocka et al., Nature, May 2006 Li et al., Nature, May 2006
Histone Marks and Their Readers Readers identified by SILAC (Stable Isotope Labeling by Amino acids in Cell culture). Vermeulen et al., Cell 2010
Chromatin Immunoprecipitation (ChIP) Biological sample Fixation des interactions Protéine-ADN au formaldehyde, et fragmentation par sonication Immuno-précipitation avec un anticorps spécifique, reconnaissant spécifiquement les modifications des queues histones En parallèle, Amplification du matériel non-spécifique (MOCK) Amplification du matériel enrichi (EXP) Hybridation des échantillons sur des bio-puces: ratio EXP/MOCK ou Séquencage Haut-débit Pour plus de détails, voir http://mescaline.igh.cnrs.fr/EpiGeneSys/www/index.php?option=com_content&view=category&layout=blog&id=62&Itemid=66
High-resolution profiling of Histone Methylations in the Human Genome Data from Barski et al., Cell 2007
H3K4me3 and H3K36me3 annotate genes and non-coding RNA The FoxP1 gene The Neat non-coding RNA Data (ChIP-seq) from Mikkelsen et al., Nature 2007
PcG and trxG proteins regulate cell memory and dynamic patterns of gene expression stem cell maintenance and plasticity proliferation differentiation PcG and trxG proteins X-chromosome inactivation cell fate determination Vertebrates Fly spermatogenesis Genomic imprinting Cancer
Opposing functions of PcG (Polycomb group) and trxG (trithorax group) complexes on chromatin trxG Nucleosome remodeling (BRM/Nurf complexes) ON H4-Ac Maintenance of active states (open chromatin) Histone acetylation and Methylation (TAC1 and ASH1 complexes) H3-K4 me3 Target gene PRE/TRE PcG OFF H3-K27 me3 Deacetylation and Methylation (ESC-E(Z) PRC2 complex) Maintenance of repressed states (compact chromatin) H2A Ub - Chromatin compaction - H2A Ubiquitination (PRC1 complex)
Methylasevs.Demethylase Polycombvs.Trithorax H3-K27me3vs.H3-K4me3 « The Ying and the Yang » on chromatin From Agger et al., Nature, August 2007
Quelques généralités • Un moyen alternatif de marquer la chromatin est l’incorparation • de variants d’histones. Des variants d’histones H2A et H3 sont connus depuis très longtemps, et ils sont aujourd’hui directement impliqués dans la spécification et le maintien des états chromatiniens. • Les histones de bases H2A, H2B, H3 et H4 sont codées par des gènes en clusters fortement exprimé lors de la phase S de réplication, tandis que les variants d’histones sont codés par des gènes séparés qui sont exprimés tout le long du cycle cellulaire. Donc, les variants d’histones peuvent être incorporés à tous moments au niveau de la chromatine.
Quelques exemples de variants d’histones: • CENP-A: la protéine H3-like des centromères chez les mammifères. • Son équivalent trouvé chez tous les eucaryotes et nommé CenH3 • H3.3: seulement 4 acides aminés différents par rapport à H3. • En dehors de la phase S, les dimers H3.3-H4 peuvent être incorporés lors de l’activation des gènes à la place des dimers H3-H4 (« variant de remplacement ») • De plus, H3.3 est enrichit 2 à 5 fois en modifications spécifiques de l’activation (acétylation de K9, K14, K18 et K23, et méthylation de K4 et K79) • Le variant H3.3 représenterait donc une marque importante des états actifs de la chromatine • H2A.Z: biochimiquement moins stable que H2A, il serait associé, tout comme H3.3, au marquage de la chromatine active • MacroH2A: marque spécifique de l’inactivation du chromosome X chez les mammifères • Pour plus d’infos: voir la revueHenikoff, Furuyama and Ahmad: « Histone variants, nucleosome, assembly and epigenetic inheritance », TRENDS in Genetics 2004, 20:320–326
La méthylation de l’ADN: modification au niveau de la molécule d’ADN, mais ce n’est pas une mutation au sens génétique du terme (ne modifie pas les appariements entre les bases nucléotidiques) • Chez les mammifères, la seule méthylation observée:5-méthylcytosine - Méthylation le plus souvent symétrique: CG GC - Les groupements CH3 sontexposésdans le grand sillon
- Les CpGs, cibles de la méthylation, se trouvent essentiellementdans les promoteurs des gènes (îlots CpG), et au niveau des séquences répétées- Il existe des ADN méthyltransferases (DNMTs humaines: DNMT1, DNMT de maintien, DNMT3a et DNMT3b, DNMT de novo) - Des répresseurs transcriptionnels lient spécifiquement l’ADN méthylé MeCP2 MBD1 MBD2
Régulations épigénétiques impliquant la méthylation de l’ADN - Certains gènes inactivés par méthylation au cours du développement (IL4), mais ce n’est pas un mécanisme prépondérant (Pb réversibilité?)- le maintien de l’inactivation du chromosome X chez les mammifères- le maintien de la répression Polycomb dépendante chez les mammifères- l’empreinte parentale (voir cours de Robert Feil)- La méthylation de l’ADN est fréquemment impliquée dans la transformation tumorale
Méthylation de l’ADN et cancer …. • De nombreuses maladies ont des déterminant génétiques • connus, qui pourraient être modifiés par des processus épigénétiques • Des régulations épigénétiques telles que la méthylation de l’ADN • pourraient être des cibles de traitements, parce qu’il semble plus facile • de modifier comment l’ADN est méthylé, que de modifier la séquence • d’ADN proprement dite • Dans la plupart des cellules cancéreuses, les ilots CpG sont hyperméthylés et une hypométhylation globale du génome est observée Ainsi, les profils de méthylation de différents gènes pourraient être des outils intéressants pour le diagnostique de certains cancers
Couplage Modif. Histones/Méthylation ADN Impliquant les protéines chromatiniennes du groupe Polycomb PRC2 (HMT) Early maintenance of repression X H3K27me PRC2 Long-term maintenance PRC1 X H2AK119ub H1K26me DNMTs PRC2 Very stable long-term maintenance PRC1 X meCpG From Bantignies and Cavalli, Current Opinion in Cell Biology, 2006
Méthylations, Polycomb et cancer …. Epigenetic stem cell signature in cancer, Nature Genetics 2006 Martin Widschwendter, Heidi Fiegl, Daniel Egle, Elisabeth Mueller-Holzner, Gilbert Spizzo, Christian Marth, Daniel J Weisenberger4, Mihaela Campan4, Joanne Young5, Ian Jacobs & Peter W Laird Abstract: Embryonic stem cells rely on Polycomb group proteins to reversibly repress genes required for differentiation. We report that stem cell Polycomb group targets are up to 12-fold more likely to have cancer-specific promoter DNA hypermethylation than non-targets, supporting a stem cell origin of cancer in which reversible gene repression is replaced by permanent silencing, locking the cell into a perpetual state of selfrenewal and thereby predisposing to subsequent malignant transformation.
For more info… http://www.igh.cnrs.fr/equip/cavalli http://www.epigenesys.eu Polycomb and cancer: http://www2.cnrs.fr/presse/communique/1672.htm Chromatin movie: http://www.dailymotion.pl/tag/nisic Website to download this course: http://www.igh.cnrs.fr/equip/cavalli/link.PolycombTeaching.html Giacomo.Cavalli@igh.cnrs.fr