Download

1 / 25
250 likes | 364 Views
A Role for Minerals in Genetic Expression. Summary:. Metal ions are not passive components of biological systems as once thought. Recent studies have shown that Fe, Cu, or Zn are capable of regulating the expression of genes that code for that metal’s storage or transport proteins.

E N D
A Role for Minerals in Genetic Expression Summary: Metal ions are not passive components of biological systems as once thought. Recent studies have shown that Fe, Cu, or Zn are capable of regulating the expression of genes that code for that metal’s storage or transport proteins
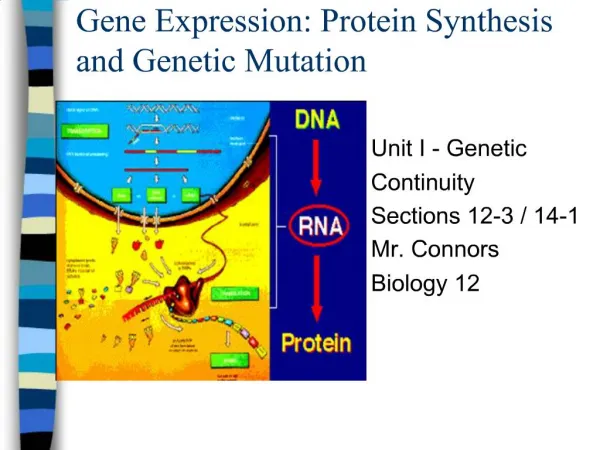
How should gene regulation be viewed DNA Level (transcription control) Transcription factors bind to the promoter region and turn on the downstream gene Base sequences (MRE’s) in promoter form the binding site The presence of the metal is needed for binding to occur RNA Level (translation control) 5’-sequences determine whether the mRNA will be read 3’-sequences stabilize mRNA to turnover Both engage proteins that are activated (deactivated) by the metal
Pre-transcription Complex
Enhancers MRP Act IIH Nucleosome TAF IIE IIF Zn Zn IIA IIB TATA TBP Pol II RNA Synthesis occurs on chromatin, not DNA. Chromatin is a protein DNA complex DNA binding domain Activation domain (recruits other proteins) Metal-responsive protein transcription factor mRNA Zn
Promoter Cyclin genes ADP P ATP Cancer of the Retina Activates E2F Cell proliferation Cyclins Stimulates CDK Inactive Cancer of the retina Rb CDK Phosphate controls cell division E2F = a transcription factor that stimulates transcription of cyclin genes Rb = retinoblastoma protein (tumor suppressor)
Early Observations that Established Metal Ion Regulation of Genes
Biochem J. 1977 Apr 15;164(1):223-8. Control of zinc-thionein synthesis in rat liver. Squibb KS, Cousins RJ, Feldman SL. The rate of [35S]cystine incorporation into hepatic zinc-thionein (a metallothionein) was stimulated after parenteral administration of 2 mg of Zn2+. Stimulation was inhibited by actinomycin D or cordycepin, supporting the concept that hepatic zinc-thionein synthesis responds quickly to changes in Zn2+ status that allow the Zn2+ to bind to the nascent thionein chains. The control of zinc-thionein synthesis by Zn2+ appears to involve changes in RNA
Inhibited by Actinomycin D X Zn induces Metallothionein MRE DNA mRNA for metallothionein Transcription factor for Zinc Zn 35S-Cystine Zn Metallothionein (binds excess cytosolic Zinc) Zn Zn
Biochem J. 1978 Feb 15;170(2):219-225. Induction of cadmium-thionein in isolated rat liver cells. Hidalgo HA, Koppa V, Bryan SE. The uptake of Cd by isolated liver cells was linearly related to the Cd concentration to which the cells were exposed. Cd-treated cells synthesized proteins de novo with the characteristics of Cd-thionein. Cycloheximide and actinomycin D prevented the synthesis of thionein. The concentration of Cd affected the amount of Cd-thionein synthesized.
Inhibited by Actinomycin D X X Cycloheximide Cd induces Metallothionein MRE DNA mRNA for metallothionein Transcription factor for Zinc Cd 35S-Cystine Cd Metallothionein (binds excess cytosolic Zinc) Cd Cd
Biochem J. 1978 Dec 1;175(3):833-40. Cell-free synthesis of metallothionein directed by rat liver polyadenylated messenger ribonucleic acid. Shapiro SG, Squibb KS, Markowitz LA, Cousins RJ. Polyribosomal metallothionein mRNA was increased at least 3-fold after of Zn2+ administration. Actinomycin D blocked the increase strongly suggesting that Zn2+ caused an elevation in metallothionein mRNA.
Transcription Start Site -60 -42 MRE 12 bp DNA Downstream Upstream Promoter Region of the metallothein gene bp = base pairs
Nucleic Acids Res. 1990 August 25; 18(16): 4683–4690. Zinc dependent binding of a liver nuclear factor to metal response element MRE-a of the mouse metallothionein-I gene and variant sequences. P F Searle Metallothionein gene transcription is inducible by zinc and other heavy metals, and several metal response elements (MREs) have been mapped within about 200 bp upstream of the site of transcription initiation in several metallothionein genes.
-200 Transcription Start Site DNA MRE-1 MRE-4 MRE-3 MRE-2 Downstream Upstream Promoter Region of the metallothein gene
Conclusions Sequences in the DNA promoter for metallothionein are specific for proteins that serve as metal-activated transcription factors Zn-specific transcription factors are activated by Zn2+, resulting in a prioritized synthesis of the mRNA coding for metallothionein protein
Proc Natl Acad Sci U S A. 1991 October 1; 88(19): 8558–8562. ACE1, a copper-dependent transcription factor, activates expression of the yeast copper, zinc superoxide dismutase gene. E B Gralla, D J Thiele, P Silar, and J S Valentine In the yeast Saccharomyces cerevisiae, copper levels exert some control over the level of SOD1 expression. We show that the ACE1 transcriptional activator protein, which is responsible for the induction of yeast metallothionein (CUP1) in response to copper, also controls the SOD1 response to copper.
Proc Natl Acad Sci U S A. 1976 Feb;73(2):371-4. Copper-induced activation of aortic lysyl oxidase in vivo. Harris ED. Day-old chicks lacking Cu had severely depressed lysyl oxidase, a Cu metalloenzyme. CuSO4 restored lysyl oxidase activity in aortic tissue. 2 hours after receiving CuSO4 (1 mg/kg) lysyl oxidase activity rose to a level 5-20 times higher than basal (saline-injected) activity. 20 hours doubled the activity. Cycloheximide suppressed activation by two-thirds. Actinomycin D caused only a 10-15% inhibition. The data suggest that Cu regulates lysyl oxidase activity and may determined its steady-state levels.
Metals Ions as Suppressors of Genetic Expression A regulatory as opposed to toxic response
Proc Natl Acad Sci U S A. 2006 Sep 12;103(37):13676-81. Epub 2006 Aug 31. NikR-operator complex structure and the mechanism of repressor activation by metal ions. Schreiter ER, Wang SC, Zamble DB, Drennan CL. Regulation of nickel concentrations in Escherichia coli is mediated by the NikR repressor via nickel-induced transcriptional repression of the nickel ABC-type transporter, NikABCDE.
Biochemistry. 2006 Mar 21;45(11):3493-505. Structural basis for the metal-selective activation of the manganese transport regulator of Bacillus subtilis. Kliegman JI, Griner SL, Helmann JD, Brennan RG, Glasfeld A. The manganese transport regulator (MntR) of Bacillus subtilis is activated by Mn(2+) to repress transcription of genes encoding transporters involved in the uptake of manganese. MntR is also strongly activated by cadmium, both in vivo and in vitro, but it is poorly activated by other metal cations, including calcium and zinc.