Download

1 / 19
260 likes | 886 Views
Induced Systemic Resistance (ISR). ISR SAR PGPR PR (1, 2, 3 etc) Acidic, basic SA INA NahG JA MeJA jar1 C2H4 ACC ein1 Priming NPR1. 1 aminocyclopropane 1 carboxylate.

E N D
ISR SAR PGPR PR (1, 2, 3 etc) Acidic, basic SA INA NahG JA MeJA jar1 C2H4 ACC ein1 Priming NPR1 1 aminocyclopropane 1 carboxylate
Acquired R reported as early as 1933, and according to Kuc, even earlier. Chester K (1933) The problem of acquired physiological immunity in plants. Quar Rev Biol 8: 129–154, 275–324. Terminology: clear distinction between SAR and ISR ca. 1992 Kloepper et al. (1992) Proposed definitions related to induced disease resistance. Biocontrol Sci. Technol 2:349-351
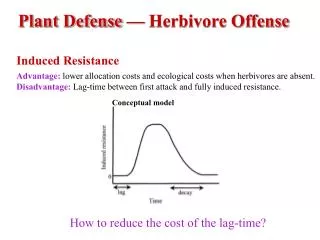
Definition and observations • ISR potentiates plant defense responses • ISR is SA-independent • ISR is independent of PR gene activation • ISR requires JA and C2H4 response pathways • ISR not associated with JA- and C2H4-responsive gene activation • ISR primes plant for enhanced C2H4 production? • Summary: compare and contrast ISR and SAR • Discussion: • 1) possible mechanisms of ISR • 2) selective advantage to plant and practical significance • Not covered in class: • 1) bacterial determinants of ISR • 2) field application of ISR
Plant-mediated*, broad-spectrum resistance response that is activated by selected strains of saprophytic rhizosphere bacteria. Many are PGPR. PGPR colonization non-specific; ability to induce SR has some specificity. * Inducing bacteria and pathogen can be spatially isolated
ISR potentiates plant defense responses • Fusarium wilt of carnation and radish • Biocontrol by P. fluorescens WCS358 • Iron competition important: sid- mutant not effective • Biocontrol by P. fluorescens WCS417 • Twice as effective • Sid-mutant still 100% effective • Worked when WCS417 and fusarium were spatially separated on the plant* • WCS417 did not trigger phytoalexin accumulation* • WCS417 treated plants produced more phytoalexin in response to Fusarium* Root rot in bean Accelerated and potentiated papilla formation *Van Peer et al (1991) Phytopathol 81:728-734
ISR is SA-independent – W Fusarium – W Pieterse et al. (1996) Plant Cell 8, 1225-1237 P. syringae
ISR is independent of PR gene activation Pieterse et al. (1996) Plant Cell 8, 1225-1237
ISR requires JA and C2H4 response pathways etr1 nahG npr1 Pieterse et al. (1998) Plant Cell 10, 1571-158
(There may be a) parallel ethylene-inducible defensive pathway that does not require NPR. A candidate pathway might be the ethylene-inducible pathway leading to Pdf1.2 gene expression that has been shown to be NPR1 independent (Penninckx et al. 1996 ). Alternatively, this low level of protection in npr1 plants may be caused by the twofold higher production of ethylene after ACC treatment (Figure 2B). However, the latter possibility seems unlikely because a twofold increase in ethylene production in wild-type Col-0 plants, by applying 2.5 mM ACC to the leaves instead of 1 mM, does not result in a higher level of protection against P. s. tomato infection (S.C.M. van Wees, unpublished results). In itself, the enhanced level of ethylene production in ACC-treated npr1 plants is intriguing because it demonstrates that npr1 plants show twofold higher ACC oxidase activity than do wild-type plants. Interestingly, pathogen infection also causes a significantly higher increase in ethylene production in npr1 plants (C.M.J. Pieterse, unpublished results), suggesting that not only SA responsiveness but also ethylene metabolism is altered by the npr1 mutation. ISR requires JA and C2H4 response pathways in that order Pieterse et al. (1998) Plant Cell 10, 1571-158
? ? Pieterse et al (2002) Plant Biology 4:535-544
But expression of this (and not others) is potentiated in plants undergoing ISR Van Wees et al. (1999) Plant Mol Biol 41:537-549 ISR not associated with JA- and C2H4-responsive gene activation So ISR is associated with potentiation of a specific set of JA-responsive genes? Also, increased sensitivity rather than increased production of JA and C2H4 (see also Pieterse et al. 2000 Physiol. Mol. Plant Pathol. 57:123-134) Pieterse et al. (1998) Plant Cell 10, 1571-158
ISR primes plant for enhanced C2H4 production? ISR plants do not show increased levels of C2H4, or JA, but ISR activated plants convert more ACC to C2H4
Pieterse et al. WORKING MODEL Pieterse (2001) Eur. J. Plant Pathol. 107:51-61
ISR vs. SAR Induced systemic resistance is induced by non-pathogenic rhizobacteria • Systemic acquired resistance is induced systemically after inoculation with necrotizing pathogens, HR, or application of some chemicals (SA analogs or agonists) Induced systemic resistance is independent of salicylic acid, but involves jasmonic acid and ethylene signaling • Systemic acquired resistance requires salicylic acid as signaling molecule in plants Induced systemic resistance is accompanied by the expression of sets of genes distinct from the PR genes • Systemic acquired resistance is accompanied by induction of pathogenesis • related proteins Both require NPR1
Discussion 1) possible mechanisms of ISR 2) selective advantage to the plant and practical significance