Download

1 / 26
260 likes | 434 Views
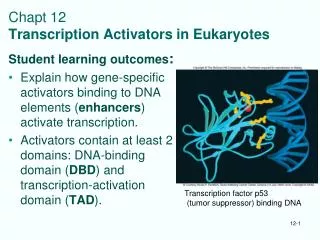
Structure and Function of Eukaryotic Transcription Activators. Many have modular structure: DNA-binding domain Transcription activating domain Proteins can have > 1 of each, and they can be in different positions in protein. Many also have a dimerization domain.

E N D
Structure and Function of Eukaryotic Transcription Activators • Many have modular structure: • DNA-binding domain • Transcription activating domain • Proteins can have > 1 of each, and they can be in different positions in protein. • Many also have a dimerization domain
Sp1: Factor for Upstream (Proximal) Class II Promoter Element • Binds GC boxes, stimulates transcription • Interacts with TAFII110 in TFIID • Also stimulates transcription of TATA-less class II promoters (by promoting TFIID binding)
Activation Domains • Acidic (e.g., GAL4, 49 aa domain – 11 acidic aa) • Glutamine-rich (e.g., 2 in Sp1, ~25% gln) • Proline-rich (e.g., CTF, 84 aa domain – 19 are proline)
DNA-binding domains • Zinc–containing motifs • Zinc fingers (Sp1 and TFIIIA) • Zinc modules (GR and other nuclear receptors) • Modules with 2 Zinc ions and 6 cysteines (GAL4) • Homeodomains - 60-aa domains originally found in homeotic mutants • bZIP and bHLH motifs - a highly basic DNA-binding domain and a dimerization domain (leucine zipper or helix-loop-helix)
Amino acid side chains in proteins can form H-bonds to DNA bases. Critical for sequence-specific binding to DNA.
3 views of C2H2 Zinc fingers Often found as repeats in a protein. Bind in the major groove of DNA.
GAL4-DNA Complex • DNA-binding domain • 2 Zn+2 bound by 6 cysteines • A Short a helix that docks into major groove Dimerization domain - Coiled coil (a helices) Fig. 12.4
Fig. 12.6 Fig 12.6
Fig. 12.7 Glucocorticoid Receptor – DNA Interactions
- Homeotic mutants have wrong organs (organ-identity mutants) - Occur in animals and plants - Important regulatory genes antennapedia Wild-type “Here’s looking at you”
12.9 • Homeotic genes are transcription factors! • Have a conserved DNA-binding domain (Homeodomain) that resembles a helix-loop-helix (HLH) domain. • Bind as a monomer
bZIP proteins • Have DNA binding and dimerization domains • DNA binding region is very basic (R and K residues) • Dimerization involves a Leucine Zipper • Can form heterodimers! Alpha helices form a coiled-coil with inter-digitating leucines
A Leucine Zipper is a Coiled Coil Motif Peptide from GCN4 Fig. 12.10
Fig 12.13 Domain Independence demonstrated with a chimeric transcription factor
Function of Activation Domains • Recruit specific components of the pre-initiation complex (a), or the holoenzyme (b).
GAL4(which binds to an upstream element) • Promotes binding of TFIIB, which promotes recruitment of the other factors and RNAP. • Probably binds directly to TFIIB (i.e., it doesn’t work by stimulating TFIID to bind TFIIB tighter) • GAL4 also promotes assembly of downstream basal factors, TFIIE and/or TFIIF+RNAP II.
Activation from a Distance: Enhancers • There are at least 4 possible models Factor binding to the enhancer induces: • supercoiling • sliding • Looping • Tracking
Fig. 12.20 Models for enhancer function
E- enhancer Psi40- rRNA promoter Transcription of DNAs 1-5 was tested in Xenopus oocytes. Results: good transcription from 2, 3, and 4 (also 2 >3 or 4) but not 5. Conclusion: Enhancer does not have to be on same DNA molecule, but must be somewhat close. Rules out the sliding and supercoiling models. From Fig. 12.22
Looping out by a prokaryotic, enhancer-binding protein visualized by EM. NtrC – protein that binds glnA enhancer and RNAP σ54 polymerase – RNAP with a 54-kDa sigma factor (defective, needs enhancer) Like Fig. 9.20
Combinatorial Transcription:expression and regulation depends on the combination of elements in the promoter human metallothionine promoter GC box MRE- metal response element BLE- enhancer that responds to activator AP1 GRE- Glucocorticoid response element Fig. 12.23
Insulators • Block enhancers • Also act as barriers to heterochromatin spreading induced by a silencer Fig 12.28
Regulation of Transcription factorsor “Regulating the Regulators” A lot of post-translational regulation: Why? - Quicker response time - Avoid silencing by keeping the transcription factor gene on (?) Some of the mechanisms: 1. Coactivators or mediators 2. Phosphorylation-dephosphorylation: can be + or - 3. Ubiquitination (deubiquitination): covalent attachment of ubiquitin (small protein) to lysines can modulate activity or trigger destruction 4. Sumoylation: covalent attachment of SUMO (small ubiquitin-like modifier) peptide to lysines, factor is inactivated but not destroyed 5. Acetylation: histoneacetyltransferases (HATs) acetylatelysines on histone and non-histone proteins, can be + or -