Download

1 / 29
290 likes | 435 Views
Radiation Biophysics Laboratory Department of Cell Biology & Biophysics , Faculty of Biology , University of Athens. Action Mechanism of Electromagnetic Fields on Cells. A pathway to Cell Death. Dr. Dimitris J. Panagopoulos Prof. Lukas H. Margaritis. Cell Membrane.

E N D
Radiation Biophysics Laboratory Department of Cell Biology & Biophysics, Faculty of Biology, University of Athens Action Mechanism of Electromagnetic Fields on Cells.A pathway to Cell Death Dr. Dimitris J. Panagopoulos Prof. Lukas H. Margaritis
Cell Membrane Cell Membranes and Ion Channels Ion Channel structure D.J.Panagopoulos, L.H.Margaritis
Cation Channels Unfolded Sodium channel protein Cross section of Na+ channel with positively charged S4 helices D.J.Panagopoulos, L.H.Margaritis
Cation Channels open by electrostatic forces on their S4 charges, generated by 30mV changes in the membrane potential Channels open by an outward rotational movement of the S4 helices Before ions pass through an open channel they enter special binding sites close to the pore. There are 3 binding sites in potassium channel close to each other and to S4 D.J.Panagopoulos, L.H.Margaritis
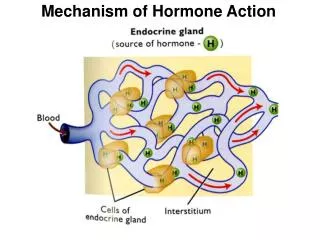
Transmembrane Electrical Potential At electrochemical balance, the net movement of ions across the membrane is zero and there is a potential differenceΔbetween the two sides, with the internal always negative relative to the external, called Transmembrane Electrical Potential Δ = o - i 100mV (s10 nm, membrane’s thickness) D.J.Panagopoulos, L.H.Margaritis
The contribution of each ion type to the total potential difference is given by the Nernst equation: o - i = - ln o,i,electrical potentials on external and internal membrane surfaces R, the Gas Constant T, the Absolute Temperature, (K) z, ionic charge (in electrons) or ion’s valence Fc, Faraday constant Co,Ci, concentrationsof the certain type of ion on the external and internal sides of the membrane at equilibrium The total transmembrane potential difference is the sum of the contributions from all kinds of free ions D.J.Panagopoulos, L.H.Margaritis
Cell’s Free Ions Transmembrane Electric Field Across all cell membranes there is a strong Electric field, (107V/m), mainly determined by free ions concentration differences between the two sides of the membrane Intensity of Transmembrane Electric Field m= Δ / s 107V/m D.J.Panagopoulos, L.H.Margaritis
Action Mechanism of EMFs on Cells Theory’s Main Points • Forces exerted by the external field on the free ions around cell membranes • Forced-Vibration induction on the free ions. • Forces on the sensors of voltage-gated channels by the oscillating ions • Irregular gating of cation channels • Disruption of cell’s electrochemical balance and function D.J.Panagopoulos, L.H.Margaritis
Any external oscillating field exerts oscillating forces on the free ions around any cell membrane inducing a forced-vibration on the whole ionic cloud A. Electric Field Consider an External Alternating E-field = o sin t =2 D.J.Panagopoulos, L.H.Margaritis
Forces exerted on each free ion Each free ion in the vicinity of a cell’s plasma membrane will accept the following forces: • External Field’s Oscillating Force: F1 = z qe= o z qe sin t, (displacementxof the ion from its initial position) z qe the ion’s charge 2.Restoration Force: F2 = - D x D = mi o2 the restoration constant, miion mass =2o, othe ion’s oscillation self frequency 3. Damping Force:F3 = -u = - 6 nf u u, ion’s vibrationall velocity λ=6nf, attenuation coefficient, nfviscosity of the medium, ion radius Calculation of the Attenuation Coefficient: a) In the aqueous solutions, (for 37oC), nf= 710-4Kg/ms, 10-10m, 10-12Kg/s. b) Within the channels from“patch-clamp” conductivity measurements (forΝa+ ions): 6.410-12Kg/s D.J.Panagopoulos, L.H.Margaritis
Forced-Vibration Equation The ion because of the above forces will acquire accelerationaand its movement equation will be: mia= - u - Dx + o z qe sin t mi + + mi o2x = o zqe sin t (1) (2nd order linear diff. eq. with constant coefficients) • All experimental data about cytosolic free ions’ spontaneous oscillations and membrane potential spontaneous oscillations, in all types of cells show that the frequencies of these oscillations are: 0.016 - 0.2 Hz • We assume that the frequencies of the spontaneous ionic oscillations represent the ionic self-frequencies, o =o/2 and we adopt the value: o = 0.1Hz D.J.Panagopoulos, L.H.Margaritis
The general solution of eq. (1), is: x = cos t + C1 e+ C2 e (2) C1,C2constants calculated from the initial conditions 1, 2roots of the “characteristic equation”: mi 2++ mio2=0 102-2.631013s-1 C1 - C2- uo410-140 We accept as maximum ion velocity: uo 0.25m/s (velocity calculated forΝa+ions, moving within the channel) D.J.Panagopoulos, L.H.Margaritis
Finally, the general solution of eq (1), is: x = cos t -(3) The constant term displaces the whole vibrational movement by: - Therefore, at the moment when the external field is applied or interrupted the ion displacement is doubled pulsed EMFs are twice more bioactive than continuous EMFs. The vibrational term:x= cos t (4) Represents an harmonic oscillation of constant amplitude, independent of any initial conditions The amplitude is: A = D.J.Panagopoulos, L.H.Margaritis
Irregular channelgating Oscillating Ions = movement of electric charge, exerting forces on the sensors of electrosensitive channels Electrosensitive channels are cation channels. They open or close by electrostatic interaction between their voltage-sensors and the transmembrane voltage. The voltage sensors of cation channels are four symmetrically arranged, transmembrane, positively charged a-helices, each one named S4. Changes in the transmembrane potential ΔΨ30mV Induce gating of the electrosensitive channels D.J.Panagopoulos, L.H.Margaritis
Displacement of one single-valence ion in the vicinity of S4 by r 10-12m, exerts on the S4 a force equal to that generated by a change of 30 mV, in the transmembrane potential: m= ΔΨ/s (5) m= F/q (6) Fthe force on anS4domain, q = 1.7qe(7) (effective charge of eachS4domain) F= q F = Δ(8) Force onS4able to gate the channel For Δ = 30mV, s = 10nm: F =8.16 10-13N(9) Force exerted onS4 by an oscillating z-valence ion: F = F = -2r r = -(10) D.J.Panagopoulos, L.H.Margaritis
ris the necessary ion displacement in theS4domain, able to gate the channel We acceptthat the distance between the oscillating ion and the effective charge of S4, is r 1nm 80(aqueous solutions) 4, (channel internal) (9), (10) r 0.810-10m, ( = 80) r 410-12m, ( = 4). A single ion’sDisplacement of 10-12min the S4 domain, can gate the channel D.J.Panagopoulos, L.H.Margaritis
An oscillating external electric field can alter cell function, if it is able to induce on the free ions a forced-vibrationof amplitude: A r(11) rdecreases proportionally to the number and charge of the ions which act simultaneously on anS4domain D.J.Panagopoulos, L.H.Margaritis
The Thermal Motion The free ions within the same time interval are displaced at longer distances because of thermal activity than because of the forced-vibration But: Thermal motion is a random motion in every possible direction, different for every ion. It does not displace the whole ionic cloud. Foru = 0.25 m/s, the time interval isδt 1.610-11s, for the necessary displacement r=10-12m. Mean Free Pathof an ion within the channel: 10-11m. The ion withinδt, will run one mean free path in the direction of the external field and tens of free paths each one in different direction because of thermal motion. Forced-vibration is a synchronized movement of the whole ionic cloud in one direction acting additively on S4. The tens of free paths because of thermal motion, each one in a different direction, result in mutually extinguishing opposing forces on the channel sensors, while at the same time one single displacement of the whole ionic cloud in phase results in additive forces on the sensor. D.J.Panagopoulos, L.H.Margaritis
EMF Bioactivity Characteristics (11)410-12m (12) o10-3(13) (Hz, V/m) logo log - 3 0(14) (one single-valence cation and continuous oscillating field) With pulsed fields, the left sides are multiplied by 2 For double-valence cations, (Ca2+), the left sides are multiplied by 4 and the right sides divided by 2 ( ris divided by 2)bioactivity is multiplied by 8. For 2 Ca2+ interacting simultaneously from 1nmdistance with anS4domain, bioactivity is multiplied by 16: 0.2510-12m (15) o0.62510-4(16) (Hz, V/m) logo log - 4.20 (17) For 3 Ca2+ interacting simultaneously from 1nmdistance with anS4domain, bioactivity is multiplied by24 D.J.Panagopoulos, L.H.Margaritis
B. Magnetic Field The mechanismis the samein the case of Oscillating Magnetic fields if we only replace the electric force F1= o zqesin t, by a B-field (B=Bosin t)forceexerted on an ion zqemoving with velocity u, vertically to the direction of the field: F1 = Bou zqe sin t (18) For one single-valence cation moving within a channel vertically to the direction of a continuous oscillating B-Fieldwithu= 0.25 m/s : 410-12m (19) (rad/s,m/s, T), Bo40 (20) (Hz, G) log Bo log +1.6(21) For 2 Ca2+ interacting simultaneously from 1nmdistance with anS4domain: 0.2510-12m (22) (rad/s,m/s, T) Bo2.5 (23) (Hz, G) log Bo log +0.4(24) D.J.Panagopoulos, L.H.Margaritis
Taking into account an induced electric fieldEind,generated by the pulsed magnetic onewith a typical intensity of 1 V/m, parallel to the magnetic forceF1, (vertically toB), for 2 Ca2+ interacting simultaneously from 1nmdistance with anS4domain: 0.2510-12m (25) (V/m, T) Bo2.5 - 4104 (26) (Hz, G) (lineB in E-B Bioactivity diagram) D.J.Panagopoulos, L.H.Margaritis
For pulsed E and B fields and 2 double-valence cations interacting simultaneously with S4 from 1nmdistance D.J.Panagopoulos, L.H.Margaritis
For frequencies 0-104Hz, even very weak E-fields of less than 1V/m, or B-fields of 1G are able to alter cell function D.J.Panagopoulos, L.H.Margaritis
According to the theory, low frequency EMFs are more bioactive than high frequency EMFs. This explains for the first time experimental data indicating that biological action of RF-Microwave fields is due to the ELF frequencies included (pulse repetition frequency, modulation) According to the theory, pulsed EMFs are more bioactive than continuous EMFs. This explains for the first time the higher bioactivity of pulsed EMFs reported in several experiments. Most RF-Microwave fields are Pulsed (GSM, DCS) or Modulated (FM) at ELF frequencies D.J.Panagopoulos, L.H.Margaritis
b S8 S4 G S3 S1 S2 S8 2a 2b G S8 FC S4 S3 S7 S2 G S1 NC OC Recent Experiments indicate DNA damage or Cell Death induction by EMFs (Panagopoulos et al., 2007; Diem et al., 2005; Markova et al., 2005; Salford et al., 2003; Lai and Singh 1995; 1996) 1 Ovariole of a sham exposed female insect with TUNEL negative egg chambers at all the developmental stages from germarium (G) to stage 8 2 Ovariole of exposed female insect with TUNEL positive signal at both check-points, germarium and stage 8 and TUNEL negative signal at the intermediate stages 3 Ovarioles of exposed female insects with TUNEL positive signals at all the developmental stages and in all types of egg chamber cells, nurse cells (NC), follicle cells (FC) and the oocyte (OC) D.J.Panagopoulos, L.H.Margaritis
Disruption of intracellular Ionic Concentrationsmay lead to Cell Death 1. Irregular increase of intracellular ion concentration, may cause water molecules to enter the cell, driven by osmotic forces, proportional to the concentration increase. This can cause the cell to swell out and the plasma membrane to get ruptured, resulting to cell necrosis. 2. Perturbations of intracellular Ca+2 concentrations are responsible for apoptotic triggering. 3. A common event leading to both apoptosis and necrosis is mitochondrial membrane permeabilization, leading to increased mitochondrial concentration of Ca+2 ions D.J.Panagopoulos, L.H.Margaritis
Conclusion Forced-Vibration of free ions in the vicinity of a cell membrane induced by external EMF can irregularly gate ion-channels, disrupt the cell’s electrochemical balance and function and may even lead to Cell Death D.J.Panagopoulos, L.H.Margaritis