Download
1 / 36
370 likes | 563 Views
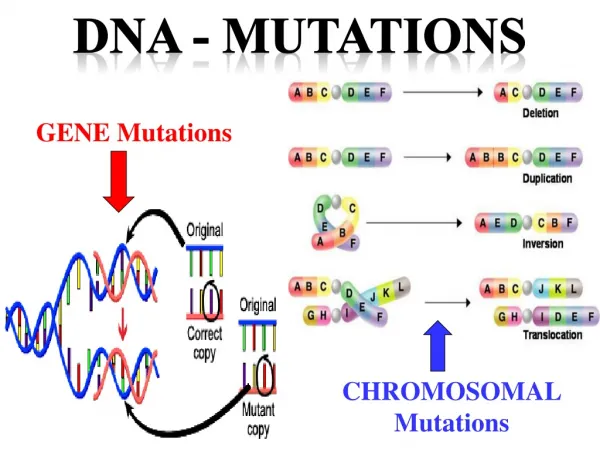
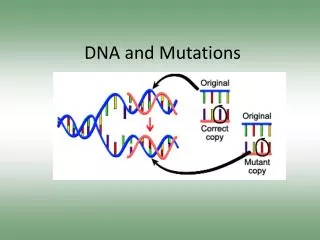
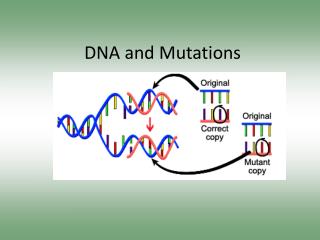
DNA Repair and Mutations. Chemical reactions and some physical processes constantly damage genomic DNA At the molecular level, damage usually involves changes in the structure of one of the strands Vast majority are corrected by repair systems using the other strand as a template

E N D
DNA Repair and Mutations Chemical reactions and some physical processes constantly damage genomic DNA At the molecular level, damage usually involves changes in the structure of one of the strands Vast majority are corrected by repair systems using the other strand as a template Some base changes escape repair and the incorrect base serves as a template in replication The daughter DNA carries a changed sequence in both strands; the DNA has been mutated Accumulation of mutations in eukaryotic cells is strongly correlated with cancer; most carcinogens are also mutagens
Molecular Mechanisms of Spontaneous Mutagenesis • Deamination • Very slow reactions • Large number of residues • The net effect is significant: 100 C ????? • U events /day in a mammalian cell • Depurination • N-glycosidic bond is hydrolyzed • Significant for purines: 10,000 purines lost/day in a mammalian cell • Cells have mechanisms to correct most of these modifications.
DNA repair enzymes • a lot of DNA damage → elevated levels of repair enzymes • extreme change in cell's environment (heat, UV, radiation) activates genes that code DNA repair enzymes • E.g., heat-shock proteins are produced in heat-shock response to high temperatures.
Damage of the double helix • Single strand damage • information is still backed up in the other strand • Double strand damage • no backup • can cause the chromosome to break up
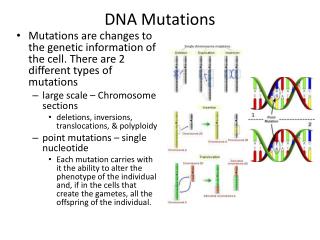
Types of DNA Repair Systems • Single strand repair • Nucleotide Excision Repair • Base Excision Repair • Mismatch Repair (shortly after replication) • Double strand repair • Homologous end-joining • Non-homologous end-joining
Types of DNA Repair Systems Mismatches arise from occasional incorporation of incorrect nucleotides Abnormal bases arise from spontaneous deamination reactions or via chemical alkylation (alk genes) Pyrimidine dimers form when DNA is exposed to UV light Backbone lesions occur from exposure to ionizing radiation
Diseases often result from failure of DNA repair systems • colon cancer • cellular ultraviolet sensitivity • Werner syndrome (premature aging, retarded growth) • Bloom syndrome (sunlight hypersensitivity)
Page 1173 Figure 30-51 Types and sites of chemical damage to which DNA is normally susceptible in vivo. Red, oxidation; blue, hydrolysis; green, methylation.
Endogenous and exogenous alkylating agents (tobacco smoke, some anticancer drugs). Alkylation destabilizes the glycoside bond and can ultimately lead to backbone breaks. O6-alkylguanine has a different pattern of H-bond donor and acceptor atoms than the parent guanine base. As a result, it base pairs with T instead of C, giving rise to G A transition after the second round of replication:
Single strand repair • Nucleotide excision repair • a large multienzyme compound scans the DNA strand for anomalies • upon detection a nuclease cuts the strand on both sides of the damage • DNA helicase removes the oligonucleotide • the gap is repaired by DNA polymerase and DNA ligase enzymes
O6-alkylguanine DNA alkyltransferase (AGT) Directly repaires alkylation damage (O6-alkylguanines) by transferring the O6-alkyl group from damaged guanine in DNA to a Cys residue in the AGT active site in a stoichiometric reaction. The protein is inactivated via alkylation and undergoes proteolytic degradation. AGT protein is highly conserved: helix-turn-helix DNA binding motif the alkylated base is “flipped” out of the helix to enter the hydrophobic alkyl-binding pocket of the protein high metabolic cost for the cell is outweighed by the need to maintain genetic integrity
Formation of pyrimidine dimers induced by UV light. (b) Formation of a cyclobutane pyrimidine dimer introduces a bend or kink into the DNA
Figure 30-52 The cyclobutylthymine dimer that forms on UV irradiation of two adjacent thymine residues on a DNA strand. Photolyase: repairs cyclobutane pyrimidine dimers. Uses the energy of light to catalyze the reversal of the cyclobutane bonds, producing intact DNA. Not very important in mammals.
Figure 30-56 Base Excision RepairDNA glycosylases hydrolyze the glycosidic bond of their corresponding altered base to yield an AP site. Page 1177
Base-excision repair pathway 1 DNA glycosylase recognizes damaged base, cleaves N-glycoside bond 2 AP endonuclease cleaves backbone near the AP site. 3 DNA pol I initiates repair synthesis from the free 3′ OH at the nick, removing and replacing the damaged strand. 4 Nick sealed by DNA ligase.
Intermediates on the base flipping pathways of Human 8-oxoguanine glycosylase hOGG1) The exo-site complex of hOGG1 with an extrahelical guanine (by chemical trapping)(left). The fully extrahelical complex with 8-oxoG is shown on the right. Note the DNA distortion in the NON substrate. Extrahelical Damaged Base Recognition by DNA Glycosylase Enzymes JamesT. Stivers Chemistry - A European Journal Volume 14, Issue 3, pages 786–793, January 18, 2008
Abstract • The efficient enzymatic detection of damaged bases concealed in the DNA double helix is an essential step during DNA repair in all cells. Emergent structural and mechanistic approaches have provided glimpses into this enigmatic molecular recognition event in several systems. A ubiquitous feature of these essential reactions is the binding of the damaged base in an extrahelical binding mode. The reaction pathway by which this remarkable extrahelical state is achieved is of great interest and even more debate.
X-Ray structure of human uracil–DNA glycosylase (UDG) in complex with a 10-bp DNA containing a U·G base pair. Mismatch flipped out. DNA helix stabilized by aa plug. Catalysis is by GAB, but activity is not abolished by loss of the acid residue in the active site. Conformational strain helps drive the reaction!! aa plug prevents base from flipping back into helix.
Base excision repair of alkylated DNA by AlkD in Bacillus cereus. Lack of + adds stability THF Resembles apurinc DNA a, AlkD catalyses the hydrolysis of the N-glycosidic bond to liberate an abasic site and free nucleobase (100X rate increase over spontaneous depurination). The enzyme is specific for positively charged N3-methyladenine (a) and N7-methylguanine (b). c, d, Structures used to trap AlkD in complex with alkylated and abasic DNA. e, Crystal structure of AlkD bound to 3d3mA-DNA. Each of the six HEAT repeats is coloured red-to-violet. The DNA is coloured silver with the 3d3mA nucleotide colored magenta. EH Rubinson et al.Nature000, 1-6 (2010) doi:10.1038/nature09428
alkD consists of HEAT repeats • HEAT: domain found in a number of cytoplasmic proteins including Huntingtin, elongation factor 3 (EF3), protein phosphatase 2A (PP2A), and the yeast PI3-kinaseTOR1and form rod-like helical structures • A substantial number of HEAT repeats have been detected in proteins involved in translation including all eIF4Gs!
Figure S3. DNA binding by the HEAT repeats of AlkD. Electrostatic surface potential (blue, positive; red, negative) of AlkD showing a high degree of positive charge within the concave DNA binding cleft. Structure based sequence alignment of HEAT repeats. Residues that contact the DNA in both substrate (3d3mA•T, G•T) and product (THF•T, THF•C) complexes are highlighted yellow, residues contacting the DNA in only substrate or only product are highlighted blue and magenta, respectively.
Crystal structures of AlkD in complex with 3d3mA-DNA (a) and THF-DNA (b). EH Rubinson et al.Nature000, 1-6 (2010) doi:10.1038/nature09428 The modified 3d3mA and tetrahydrofuran (THF) nucleotides are colored blue, and the opposing thymine is magenta.
DNA positioned along +charged concave surface of alkD Contacts 10 bp Contacts cluster around mismatch Fewer contacts on lesion strand
Both 3m3A and THF reside on the face of the DNA duplex NOT in contact with the protein whereas the compelement is nestled in the enzyme cleft. This THF trapped complex shows the abasic site rotated 90o around the PD backbone where it is totally solvent exposed!! Opposing T has slipped out of helix into DNA minor groove!! Backbone distorted NO aa PLUG DNA duplex has collapsed to retain stacking interactions.
Recognition of DNA damage by AlkD. a, 3d3mA-DNA (substrate) complex. b, THF-DNA (product) complex. Dashed arrows denote displacement of THF and opposing thymine from their positions in B-DNA. Hydrogen bonds are shown as dashed lines. Views are down the DNA helix axis.
Remodelling of a G•T wobble base pair by AlkD. a, AlkD–G•T-DNA complex viewed down the helical axis. b, The structure of a G•T wobble base pair in DNA alone is superimposed onto the AlkD–G•T complex. Steric clashes between the protein and DNA are highlighted by yellow stars, and disrupted hydrogen bonds (dashed lines) are shown by a red X. c, Relative single-turnover rates (kst) of 7mG excision from a 25mer oligonucleotide duplex by wild-type AlkD and the indicated AlkD mutants. AlkD seems to detect DNA duplex destabilization rather than specifically recognizing modified bases. Protein restructures the wobble and disrupts base stacking.
Solvent exposure increases the lifetime that spontaneous depurination is likely to occur!!!The phosphate groups on the DNA may participate in the rate enhancement by positioning water molecules for solvent attack on the glycoside bond.
AlkD captures the DNA in an orientation that holds the orphaned base next to the protein and exposes the lesion to a hydrolytic environment. The distorted DNA conformation is stabilized not by a side chain plug, but by stacking of flanking base pairs as a result of both lesion and orphaned base flipping. Phosphate (gold “P”) assisted hydrolysis could occur either by positioning of the water molecules adjacent to the C1' carbon in a dissociative hydrolysis reaction, or through stabilization of an oxocarbenium ion intermediate Figure S12. Proposed mechanism for how AlkD facilitates hydrolysis of N3 and N7-alkylpurines by distorting the DNA backbone.
Methylation and mismatch repair Really only understood well in E.coli. The methylation occurs at the N6 of adenines in (5′)GATC sequences. (palindrome) Dam=DNA adenine methylation
Double strand repair • Homologous end-joining • damaged site is copied from the other chromosome by special recombination proteins
Double strand repair • Nonhomologous end-joining • only in emergency situations • two broken ends of DNA are joined together • a couple of nucleotides are cut from both of the strands • ligase joins the strands together
Cell Cycle and DNA repair • Cell cycle is delayed if there is a lot of DNA damage. • Repairing DNA as well as signals sent by damaged DNA delays progression of cell cycle. This ensures that DNA damages are repaired before the cell divides