Download

1 / 59
610 likes | 759 Views
CHAPTER 24 Genes and Chromosomes. Organization of information in chromosomes DNA supercoiling Structure of the chromosome. Key topics : . Management and Expression of Genetic Information. Previous chapters dealt with

E N D
CHAPTER 24Genes and Chromosomes Organization of information in chromosomes DNA supercoiling Structure of the chromosome Key topics:
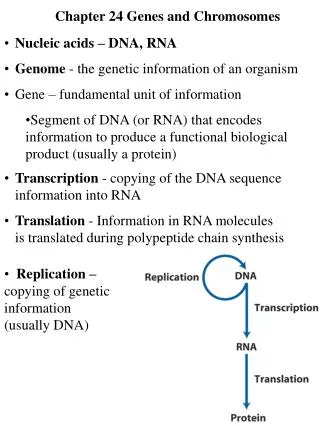
Management and Expression of Genetic Information Previous chapters dealt with metabolic pathways, in which the chemical structures of small molecules were modified by enzymes signal transduction pathways, in which interactions of ligands with receptor proteins caused physiological responses The following chapters deal with information pathways, in which genetic information stored as the nucleotide sequence is maintained and expressed
The Central Dogma of Molecular Biology The discovery of double-helical structure of DNA in 1953 laid a foundation to thinking of biomolecules as carriers of information It was well understood by 1950 that proteins play roles of catalysts but their role in information transfer was unclear Francis Crick proposed in 1956 that “Once information has got into a protein it can’t get out again” The Central Dogma was proposed by Francis Crick at the time when there was little evidence to support it, hence the “dogma”
How does genes function? Central Dogma: DNA to RNA to Protein.
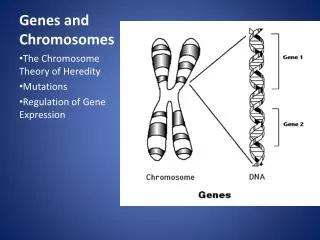
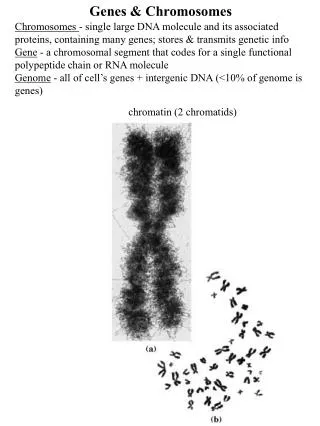
Genes and Chromosomes • What is gene? • One gene-one enzyme. • One gene-one protein (polypeptide). • Genes are segments of DNA that code for polypeptides and RNAs. • What is chromosome? Chromosome consists of one covalently connected DNA molecule and associated proteins • Viral genomic DNA may be associated with capsid proteins • Prokaryotic DNA is associated with proteins in the nucleoid • Eukaryotic DNA is organized with proteins into a complex called the chromatin
DNA is a Very Large Macromolecule The linear dimensions of DNA are much bigger than the virions or cells that contain them Bacteriophages T2 and T4 are about 0.2 m long and 0.1 m wide Fully extended T4 DNA double helix is about 60 m long DNA in the virion or cell is organized into compact forms, typically via coiling and association with proteins
The Size and Sequence of DNA Molecules in Bacteria and their viruses Bacteria(E. coli) 4,639,221 1.7 mm 0.002 mm
DNA, Chromosomes, Genes, and Complexity Note that despite the trends in the previous table, neither the total length of DNA, nor the number of chromosomes correlates strongly with the perceived complexity of the organisms Amphibians have much more DNA than humans Dogs and coyotes have 78 chromosomes in the diploid cell Plants have more genes than humans The correlation between complexity and genome size is poor because most of eukaryotic DNA is non-coding Recent experimental work by Craig Venter suggests that a minimal living organisms could get by with less than 400 genes
Eukaryotic genomes have several sequence components • Nonrepetitive DNA: the complexity of the slow component corresponds with its physical size, i.e., unique sequences. • Moderately repetitive DNA:.component with a Cot1/2of 10-2 and that of nonrepetitive DNA. Contains families of sequences that are not exactly the same, but are related. The complexity is made up of a variety of individual sequences, each much shorter, whose total length together comes to the putative complexity. Usually dispersed throughout the genome. • Highly repetitive DNA: component which reassociates before a Cot1/2of 10-2. Usually forms discrete clusters.
Composition of the Human Genome Notice that only a small fraction (1.5 %) of the total genome encodes for proteins The biological significance of non-coding sequences is not all clear Some DNA regions directly participate in the regulation of gene expression (promoters, termination signals, etc) Some DNA encodes for small regulatory RNA with poorly understood functions Some DNA may be junk (pieces of unwanted genes, remnants of viral infections
Many eukaryotic genes contain intervening sequences (introns)
Some Bacterial Genomes Also Contain Introns It was thought until 1993 that introns are exclusive feature of eukaryotic genes About 25% of sequenced bacterial genomes show presence of introns Introns in bacterial chromosome do not interrupt protein-coding sequences; they interrupt mainly tRNA sequences Introns in phage genomes within bacteria interrupt protein-coding sequences Many bacterial introns encode for catalytic RNA molecules that have ability to insert and reverse transcribe themselves into the genomic DNA
Transposons DNA sequence is not completely static Some sequences, called transposons, can move around within the genome of a single cell The ends of transposons contain terminal repeats that hybridize with the complementary regions of the target DNA during insertion To be covered in Ch. 25.
Important Structural Elements of the Eukaryotic Chromosome Telomeres cap the ends of linear chromosomes and are needed for successful cell division Centromere functions in cell division; that’s where the two daughter chromosomes are held together during mitosis (i.e. after DNA replication but before cell division)
Centromere: Mitotic segregation of chromosomes. Simple-sequence DNA is located at centromere in higher eukaryotes. Telomere: At ends of chromosomes. (TTAGGG)n in human.
Telomeres and Cellular Aging In many tissues, telomeres are shortened after each round of replication (end-replication problem of linear DNA); the cellular DNA ages Normal human cells divide about 52 times before losing ability to divide again (Hayflick limit)
How is DNA packed in the chromosomes • DNA Supercoiling. • Proteins assisted packaging (nucleosomes)
DNA Supercoiling DNA in the cell must be organized to allow: Packing of large DNA molecules within the cells Access of proteins to read the information in DNA sequence There are several levels of organization, one of which is the supercoiling of the double-stranded DNA helix
Supercoiling of DNA can only occur in closed-circular DNA or linear DNA where the ends are fixed. Underwinding produces negative supercoils, wheres overwinding produces positive supercoils.
Negative and positive supercoils . Topoisomerases catalyze changes in the linking number of DNA.
Supercoiling induced by separating the strands of duplex DNA (eg., during DNA replication)
Negative supercoils facilitate separation of DNA strands (may facilitate transcription)
Topology of cccDNA is defined by: Lk = Tw + Wr, where Lk is the linking number, Tw is twist and Wr is writhe.
Intertwining of the two strands • Nodes = ss crossing on 2D projection. Right-handed crossing = +1/2 Left-handed crossing = -1/2 Lk = number of times one strand winds around the other on 2D projection. One linking number = 2 nodes.
Topoisomerases are Targets for Antibiotics and Anti-cancer Drugs Bacterial topoisomerase inhibitors Type I topoisomerase inhibitors
DNA damages are produced by topoisomerase inhibitors • Most topoisomerase inhibitors act by blocking the last step of the topoisomerase reaction, the resealing of the DNA strand breaks. Therefore, these inhibitors will produce single-strand or double-strand DNA breaks in the DNA.
DNA Compaction Requires Solenoidal Supercoiling, not plectonemic supercoiling.
Protein-assisted Packaging of DNA Nucleosomes are the fundamental organizational units of eukaryotic chromatin
Each nucleosome has a histone core wrapped by DNA (146 bps) in a left-handed solenoidal supercoil about 1.8 times. The linker DNA is about 54 bps in length.