Download
1 / 67
690 likes | 1.01k Views
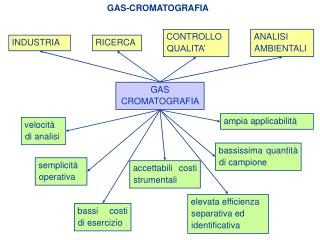
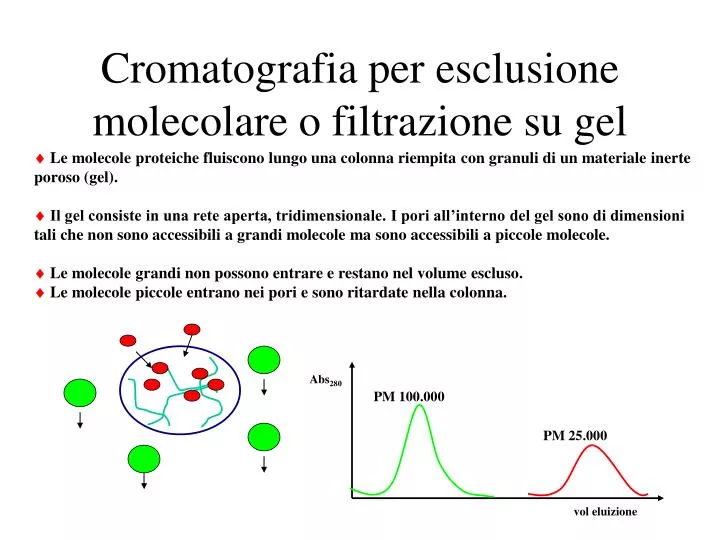
Cromatografia per esclusione molecolare o filtrazione su gel. Le molecole proteiche fluiscono lungo una colonna riempita con granuli di un materiale inerte poroso (gel). Il gel consiste in una rete aperta, tridimensionale. I pori all’interno del gel sono di dimensioni

E N D
Cromatografia per esclusione molecolare o filtrazione su gel Le molecole proteiche fluiscono lungo una colonna riempita con granuli di un materiale inerte poroso (gel). Il gel consiste in una rete aperta, tridimensionale. I pori all’interno del gel sono di dimensioni tali che non sono accessibili a grandi molecole ma sono accessibili a piccole molecole. Le molecole grandi non possono entrare e restano nel volume escluso. Le molecole piccole entrano nei pori e sono ritardate nella colonna. Abs280 PM 100.000 PM 25.000 vol eluizione
Il volume morto (V0) (detto anche volume della fase mobile) corrisponde al volume della colonna non occupato dalla fase stazionaria solida.
Gel filtrazione: condizioni Il materiale di supporto dovrebbe essere completamente inerte rispetto alla proteina Adsorbimento di proteine sul gel Interazione elettrostatica – si evita utilizzando alta f. ionica Interazione idrofobica – si evita utilizzando bassa f. ionica La capacità risolutiva dipende dal rapporto volume colonna/volume campione, dal grado di diffusione del campione e da un eventuale comportamento non ideale della colonna. Omogeneità del campione importante perché: Inconveniente: Effetti gravitazionali fanno sì che le parti più dense del campione penetrino nel gel producendo un fronte diffuso. Si può ovviare a ciò facendo correre la colonna al contrario
Gel filtrazione: condizioni Turbolenza del flusso può causare allargamento della banda proteica Si può ovviare a ciò diminuendo la velocità di flusso. Granuli fini possono migliorare la qualità della separazione migliore risoluzione raggiungimento più veloce dell’equilibrio Inconvenienti dei granuli fini: necessitano maggior pressione di esercizio con conseguente possibile distorsione e blocco del flusso Granuli fini e rigidi
Considerazioni pratiche Volume campione = 1 – 3 % volume colonna Forma della colonna colonna larga colonna lunga vantaggi: maggior vel. di flusso svantaggi: minor risoluzione, difficoltà ad applicareil campione in maniera omogenea vantaggi: piccole distorsioni orizzontali nell’ applicazione del campione non provocano grosse perdite di risoluzione svantaggi: minor velocità di flusso Altezza colonna: 20 – 40 volte il diametro Il gel viene risospeso nella soluzione tampone e versato nella colonna in un singolo step. Vel. di flusso – dipende dal tipo di gel Gel costituito da particelle soffici: 2-5 cm/h (destrano (Sephadex); poliacrilammide (Biogel)) Gel costituito da particelle rigide: 10-15 cm/h (destrano/poliacrilammide (Ultrogel) Destrano/bisacrilammide (Sephacryl))
Applicazione e raccolta del campione Applicazione campione volume campione: 1-3% vol. colonna, conc: 10-20 mg/ml Raccolta campione
Ve è funzione approssimativamente lineare del log di massa molecolare
nucleotidi Acidi nucleici Aminoacidi proteine ELETTROFORESI MOLECOLE CHE POSSIEDONO GRUPPI IONIZZABILI Metodo di separazione basato sulla diversa velocità di migrazione di particelle cariche sotto l’influenza di un campo elettrico.
ELETTROFORESI Migrazione di particelle cariche sotto l’azione di un campo elettrico. Tecnica soprattutto ANALITICA ma anche PREPARATIVA. E’ un mezzo di separazione molto potente, fra i piu’ usati in biochimica • Metodi FRONTALI - in soluzione libera • Metodi ZONALI – attraverso un mezzo poroso Acetato di cellulosa gel carta
UN METODO ANALITICO PER CAPIRE: • NUMERO di proteine presenti in una miscela • GRADO DI PUREZZA • PUNTO ISOELETTRICO • PESO MOLECOLARE
Elettroforesi:is a method whereby charged molecules in solution, proteins and nucleic acids, migrate in response to an electrical field Una molecola proteica, a pH I.E.P, ha carica totale 0 Their rate of migration through the electrical field, depends on the strength of the field, on the net charge, size, and shape of the molecules, and also on the ionic strength, viscosity, and temperature of the medium in which the molecules are moving SI MUOVE IN UN CAMPO ELETTRICO allo stato stazionario Ez = 6 r v • E = campo elettrico • z = carica • = viscosità del mezzo r = raggio (raggio di Stockes) v = velocità La mobilità elettroforetica u = v/E = z/6 r, è proporzionale al rapporto carica/raggio In soluzione è possibile separare le molecole PM 20.000 carica –5 PM 40.000 carica –5 Hanno diverso rapporto carica/raggio A parità di carica, la mobilità elettroforetica u log PM
Fattori che influenzano la velocità di migrazione: 1. CAMPIONE (Carica, Dimensioni, Forma) 2. TAMPONE (Concentrazione, pH) 3. SUPPORTO (setacciante, non setacciante) SUPPORTI Non setaccianti (Carta), acetato di cellulosa Setaccianti Gel di poliacrilammide (PAG) Gel di Agarosio
Setaccio molecolare un gel in cui la resistenza al movimento di una particella aumenta all’aumentare delle dimensioni della particella stessa. In un gel con capacità di setaccio molecolare aumenta la differenza di mobilità tra proteine di diverso peso molecolare.
Agarose and Polyacrylamide • Although agarose and polyacrylamide differ greatly in their physical and chemical structures, they both make porous gels. • A porous gel acts as a sieve by retarding or, in some cases, by completely obstructing the movement of macromolecules while allowing smaller molecules to migrate freely. • By preparing a gel with a restrictive pore size, the operator can take advantage of molecular size differences among proteins
Agarose and Polyacrylamide • Because the pores of an agarose gel are large, agarose is used to separate macromolecules such as nucleic acids, large proteins and protein complexes • Polyacrylamide, which makes a small pore gel, is used to separate most proteins and small oligonucleotides. • Both are relatively electrically neutral
Other Roles of the Solid Support Matrix? It inhibits convection and diffusion, which would otherwise impede separation of molecules It allows a permanent record of results through staining after run.
The Net Charge is Determined by the pH of the Medium • Proteins are amphoteric compounds, that is, they contain both acidic and basic residues • Each protein has its own characteristic charge properties depending on the number and kinds of amino acids carrying amino or carboxyl groups • Nucleic acids, unlike proteins, are not amphoteric. They remain negative at any pH used for electrophoresis
Elettroforesi su gel Il gel è costituito da una rete di filamenti che formano pori e quindi molecole con carica uguale ma con dimensioni diversesi separano per effetto di forze frizionali diverse. Il gel è costituito da acrilammide (CH2=CH-CO-NH2) In presenza di (NH4)2S2O8 e di TEMED (N,N,N’,N’-tetrametiletildiammina) l’acrilammide da’ luogo a polimeri. Queste unità formano il polimero. Se è presente anche N,N’-metilene bis-acrilammide anche essa partecipa alla polimerizzazione formando legami tra le catene
Reazione esotermica (DH = -84 kJ/mol) Il radicale persolfato funge da precursore di molecole radicaliche, le quali agiscono come “iniziatori di catena” reagendo con l’acrilamide
Polimero La quantità di iniziatore influenza la struttura del gel finale: 1. Un eccesso causa un decremento della lunghezza media delle catene e quindi perdita di elasticità 2. Un difetto produce reazione troppo lenta e quindi aumenta la probabilità che l’O2 atmosferico agisca da terminatore di catena
Concentrazione e separazione • La porosità del gel dipende da • Lunghezza catene (dipende da conc. acrilammide) • Numero di legami tra le catene (dipende dal rapporto tra • acrilammide e bis-acrilammide) Conc. acrilammide (%) Range di separazione 15 10 7.5 5.0 12000 – 43000 16000 – 68000 36000 – 94000 57000 - 212000
SDS-PAGE Le proteine vengono fatte reagire con un detergente anionico, sodio dodecilsolfato (H3C-(CH2)10-CH2OSO3- Na+e formano complessi carichi negativamente. 1.4 g di SDS legano circa 1.0 g di proteina. La quantità di SDS che si lega è proporzionale al PM della proteina e quindi anche la carica negativa che questa assume è proporzionale al suo PM, ma hanno uguale densità di carica o carica per unità di lunghezza. Qualsiasi sia il punto isolelettrico delle molecole proteiche esse diventano cariche negativamente e possono essere separate su gel in base alla carica negativa acquisita dipendente dal suo PM - - - - - - proteina - - - - - - - SDS viene utilizzato insieme ad agenti riducenti (DTT) che rompono ponti S-S, trattando a caldo il campione. Tale trattamento permette di dissociare le molecole nelle loro subunità.
RUOLO DELL’SDS NELLA CORSA LE CARICHE, IL PM E LA FORMA DI UNA PROTEINA NATIVA INFLUISCONO SULLA CORSA PROTEINA NATIVA L’SDS LEGA LE PROTEINE IN UN RAPPORTO DI UN ANIONE OGNI 2 aa PERDITA DELLE STRUTTURE SECONDARIA E TERZIARIA ACQUISIZIONE DI UNA CARICA NETTA NEGATIVA RAPPORTO CARICA/MASSA SIMILE PER TUTTE LE PROTEINE NEL CAMPIONE Il trattamento con SDS elimina le differenze di forma, quindi la lunghezza della catena polipeptidica, che riflette la massa, e’ l’unico determinante della velocita’ di migrazione. La separazione avviene quindi per differenza fra pesi molecolari visto che il rapporto massa carica per ogni proteina denaturata con SDS rimane costante.
Continuous and Discontinuous Buffer Systems • A continuous system has only a single separating gel and uses the same buffer in thetanks and the gel • In a discontinuous system a nonrestrictive large pore gel, called a stacking gel, is layered on top of a separating gel • The resolution obtainable in a discontinuous system is much greater than that obtainable in a continuous one. However, the continuous system is a little easier to set up
stacking gel: è la parte superiore del gel e la sua funzione è quella di concentrare il campione proteico caricato negli appositi pozzetti, in modo che tutti i campioni comincino la loro migrazione dallo stesso punto di partenza. running gel: è la parte inferiore e la sua funzione è quella di separare le proteine dei vari campioni sulla base del loro peso molecolare. È composto dagli stessi ingredienti dello stacking gel, ma in quantità diverse. In particolare è la concentrazione di acrilammide a variare, a seconda della porosità desiderata: concentrazioni maggiori portano a pori di dimensioni minori, dunque capaci di separare le proteine con una risoluzione maggiore.
Gel su lastra 0.5 – 1 mm spessore Tris Cl pH 6.8 Tris Cl pH 8.8 Tris(idrossimetil)amminometano
IL GEL VA IMMERSO IN UNA SOLUZIONE DI CORSA CONTENENTE GLICINA 25 mM Tris HCl pH 8.3 192 mM Glicina 0.1% SDS
Gel discontinuo La concentrazione (o impaccamento) delle proteine in una banda sottile viene ottenuta grazie al pH discontinuo Infatti nei due gel sono presenti tamponi (Tris-HCl) a pH diverso (stacking pH 6.8 e running pH 8.8) Nel tampone di corsa (Tris-Glicina pH 8.3) è presente anche un diverso tipo di ioni (Glicinato) Anche il campione è tamponato a pH 6.8 nel loading buffer
Arrivata al loading buffer e allo stacking (entrambi a pH 6.8) la glicina è quasi neutra ed è la specie più lenta, mentre gli ioni Cl-sono la specie più veloce Nel buffer a pH 8.3 la glicina è carica negativamente e comincia a migrare con una certa velocità Gly Gly pH 8.3 Gly Gly Cl- Cl- pH 6.8 Cl- Cl- Cl- Cl- pH 8.8 Cl- Cl- Cl- Questo fa sì che le proteine vengano racchiuse in una regione ad elevata mobilità elettroforetica
EFFETTO STACKING Gly PROTEINE COLORANTE 6.8 Cl- Cl- Cl- Cl- 8.8 All’inizio del separating gel il pH 8.8 riporta la glicina ad un rapporto carica/massa maggiore rispetto a quello delle proteine. La glicina supera i polipeptidi, che non trovandosi più in una regione ad elevata mobilità elettroforetica rallentano notevolmente e si ritrovano compattati in una linea molto sottile. Ora le proteine si trovano tutte allo stesso livello. Nel separating la concentrazione di acrilamide è più alta, e comincia la separazione del campione in base al peso molecolare.
Gel discontinuo Il gel SDS-PAGE può essere effettuato con un sistema tampone discontinuo ad un pH diverso (pH 8.3) da quello che costituisce il gel, a sua volta composto di 2 fasi a pH diverso. I campioni vengono concentrati in zone molto piccole, portando ad una migliore separazione delle diverse specie Il sistema è costituito da uno stacking gel (pH 6.8) e da un resolving gel (pH 8.8). Il campione viene introdotto nello stacking gel. In questa zona, sotto l’effetto del campo elettrico, gli ioni si muovono; i complessi proteina-SDS, hanno mobilità intermedia tra gli ioni Cl- e la glicina presente nel tampone di corsa. Quando le specie raggiungono il resolving gel a pH 8.8, la glicina, completamente ionizzata supera i complessi proteina-SDS. Questi quindi si separano in un campo elettrico costante sulla base della loro carica, all’interno del resolving gel.
La mobilità di una proteina è direttamente proporzionale al logaritmo decimale della MASSA MOLECOLARE
Determining Molecular Weights of Proteins by SDS-PAGE • Run a gel with standard proteins of known molecular weights along with the polypeptide to be characterized • A linear relationship exists between the log10 of the molecular weight of a polypeptide and its Rf • Rf = ratio of the distance migrated by the molecule to that migrated by a marker dye-front • The Rf of the polypeptide to be characterized is determined in the same way, and the log10 of its molecular weight is read directly from the standard curve
NATIVE-PAGE Si fa in assenza di SDS. Mentre nel SDS-page la mobilità dipende principalmente dal PM, nel native-page la mobilità dipende dalla carica e dal dimensione idrodinamica della proteina. Quindi dipende dalla specifica composizione amminoacidica della proteina, pH del running buffer. Dato che la proteina mantiene una struttura foldata, la sua mobilità varia con la natura della sua conformazione (più alta mobilità per conformazioni più compatte, più bassa per strutture più grosse come oligomeri e proteine unfoldate). Quindi native-page sono sensibili a qualunque processo che altera la carica o la conformazione della proteina. Questo lo rende un ottimo strumento per studiare: • Cambiamenti della carica a causa di degradazione chimica (es. deamminazione) • Conformazioni proteiche folded, unfolded o molten globule • oligomeri e aggregati (sia covalenti che non covalenti) • fenomeni di legame quali proteina-proteina o proteina-leganti
Preparazione del gel SDS di acrilammide Resolving gel (15%) (per 2 minigel) H2O 2.5 ml Lower Tris pH 8.8* 2.5 ml Acrilammide-bis metilene acrilammide (30-08)** 5.0 ml 10% APS*** 35 l TEMED**** 6 l *18.17 g Tris + 0.4 g SDS in 100 ml soluzione. Portare a pH 8.8 con HCl ** Soluzione acquosa contenente 30% acrilammide e 0.8% bis-metile acrilammide ***Genera le specie radicaliche che permettono la polimerizzazione ****TEMED = N,N,N’,N’-tetrametilen-diammina. Catalizza la reazione di formazione di radicali da parte di (NH4)2S2O8. Stacking gel (5%) (dose per 2 minigel) H2O 2.9 ml Upper Tris pH 6.8* 1.25 ml Acrilammide-bis metilene acrilammide (30-08)** 835 l 10% APS 20 l TEMED 8 l *6.6 g Tris + 0.4 g SDS in 100 ml soluzione. Portare a pH 6.8 con HCl
Trattamento campioni Trattamento campioni 14 l di ogni campione vengono scaldati a 100 °C per 2’ insieme a 7 l di una soluzione (3xFSB) composta da: 150 ml Tris-HCl pH 6.8 150 mM DTT 6% SDS 0.3 % blue di bromofenolo 30% glicerolo Bollire a b.m. tutti i campioni per 2’. Centrifugare e caricare su gel 10 l .
Corsa e sviluppo del gel Soluzione tampone per la corsa elettroforetica 25 mM Tris 250 mM glicina 0.1 % SDS pH 8.3 Voltaggio 15 V/cm Sviluppo del gel Immergere il gel per 30’ – 12 ore in una soluzione contenente 0.25 g di Coomassie Brilliant Blue, 45 ml CH3OH, 10 ml CH3COOH, 45 ml H2O. Questa soluzione è in grado di complessarsi alle catene proteiche in modo aspecifico mediante i gruppi cationici (ARG preferezialmente) presenti sulla loro superficie. Rimuovere l’eccesso di colorante immergendo il gel in una soluzione contenete 30% CH3OH, 10% CH3COOH, 60% H2O.
Alla fine della corsa il gel può essere colorato con blu Coomassie. Il colorante lega arginina, lisine e istidine, rendendo visibili le proteine su gel in base alla loro quantità. Le intensità non sono quantitative se le proteine sono a PM molto diverso !