Download

1 / 45
610 likes | 1.56k Views
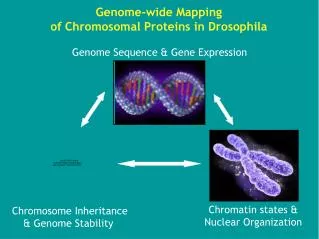
개요 : 진핵세포의 유전체는 어떻게 작용하며 진화하는가. 인간 유전체는 약 25,000 개의 유전자가 있음 . 이는 원핵세포의 5 배 정도 인간 유전체에는 RNA 난 단백질의 정보를 갖지 않는 DNA 부위를 엄청난 양으로 갖고 있어 , 질서정연하게 정리될 필요가 있음 진핵세포에서는 염색질 (chromatin) 이라는 DNA- 단백질 복합체가 고도로 정돈된 구조로 정렬 그림 19.1 : 발생 중인 도롱뇽 난자의 진핵 염색체의 DNA. 그림 19.1 발생 중인 도롱뇽 난자의 진핵 염색체의 DNA.

E N D
개요: 진핵세포의 유전체는 어떻게 작용하며 진화하는가 • 인간 유전체는 약 25,000개의 유전자가 있음. 이는 원핵세포의 5배 정도 • 인간 유전체에는 RNA난 단백질의 정보를 갖지 않는 DNA 부위를 엄청난 양으로 갖고 있어, 질서정연하게 정리될 필요가 있음 • 진핵세포에서는 염색질(chromatin)이라는 DNA-단백질 복합체가 고도로 정돈된 구조로 정렬 • 그림 19.1: 발생 중인 도롱뇽 난자의 진핵 염색체의 DNA
그림 19.1 발생 중인 도롱뇽 난자의 진핵 염색체의 DNA
개념 19.1: 염색질 구조는 일련의 DNA 응축과정에 의해 만들어진다 • 진핵 염색체는 짧은 길이에 비해 엄청난 양의 DNA를 가짐 • 사람의 경우, 각 염색체는 한 분자의 이중나선 DNA를 가지며, 평균 약 1.5 x 108 bp이며, 약 4 cm 정도의 길이 뉴클레오좀 또는 “실에 감긴 구슬”(Nucleosomes, or “Beads on a String”) • 염색질 내의 히스톤(histone) 단백질의 양은 DNA와 비슷 • 히스톤의 아미노산들에는 양성 전하를 띠는 것(아르지닌과 리신 등)이 많아 음성 전하의 DNA와 강하게 결합 • 염색질은 그림 19.2a처럼 구슬이 실에 꿴 것처럼 보임(10-nm 섬유). 이 구슬은 뉴클레오좀(nucleosome)이며, DNA 포장의 기본 단위임. 구슬 사이를 링커(linker) DNA라 부름 • 하나의 뉴클레오좀은 4개의 히스톤 단백질인 H2A, H2B, H3, H4가 각각 2개씩 있는 단백질 복합체에 DNA가 감겨 있는 모양 • 각 히스톤 단백질의 아미노말단(N-말단)은바깥쪽으로 뻗어 있어 히스톤 꼬리라 부름 DNA 응축의 다음 단계(Higher Levels of DNA Packing) • 히트톤 H1의 도움으로 30-nm 섬유 형성(그림 19.2b) • 비히스톤 단백질의 골격에 부착되어 300-nm 섬유 형성(그림 19.2c) • 염색체의모양 (그림 19.2d) • 이질염색질(heterochromatin)과 진정염색질(euchromatin)
개념 19.2: 유전자 조절은 여러 단계에서 가능하지만, 주된 조절은 전사 단계에서 이루어진다 • 모든 생명체는 항상 어떤 유전자를 발현할 것인지를 조절(외/내부 신호에 반응) • 다세포생물이 발생을 하는 동안 세포분화(celldifferentiation) 과정을 겪는데, 이때 유전자 발현은 장기적으로 조절 받음 • 사람의 경우 200여 종의 분화된 세포 유형이 있음 차별적 유전자 발현(Differential Gene Expression) • 사람의 경우, 특정 세포는 전체 유전자의 약 20% 정도를 특정 시간과 장소에서 발현. 고도로 분화된 세포(예, 근육 세포)는 더 적은 수의 유전자를 발현. 즉, 한 개체 내에서 세포 유형의 차이는 유전자의 차이가 아니라 유전자 발현의 차이이며, 이를 차별적유전자발현(differentialgeneexpression)이라 함 • 그림 19.3: 진핵세포에서 유전자가 발현되는 전 과정을 요약 • 모든 생명체에서 유전자 발현 조절의 가장 주된 지점은 전사단계
염색질 구조의 조절(Regulation of Chromatin Structure) • 염색질구조는 DNA를 핵 속에 압축해서 넣어 둘 수 있게 할 뿐 아니라 유전자 발현 조절에도 중요한 역할을 함 히스톤 단백질의 변형(Histone Modifications) • 히스톤 단백질의 변형(modification)이 유전자 전사에 영향을 준다는 것은 이미 기정사실 • 히스톤 단백질의 N-말단은 뉴클레오좀 바깥으로 노출되어 있는데(그림 19.4a), 이들 히스톤 꼬리가 다양한 변형 효소들과의 연결고리 역할 • 히스톤 단백질의 아세틸화 반응(histoneacetylation)은아세틸기(-COCH3)가 히스톤 꼬리 부위의 리신 아미노산에 결합하는 반응으로 양성전하를 잃게 되어 뉴클레오좀 간의 연결이 느슨해지게 하여 유전자의 전사를 용이하게 함(그림 19.4b). 즉, 히스톤 아세틸화 효소는 염색질 구조를 변화시키거나 다른 전사인자들을 가까이에 불러 모아 유전자의 전사를 촉진시킬 수 있음 • 다른 종류의 화학기가 가역적으로 히스톤 단백질의 꼬리에 붙을 수 있는데, 예를 들어 메틸기(methyl group, -CH3)가 붙으면 염색질의 응축을 가져옴.
DNA 메틸화 반응(DNA Methylation) • DNA가 합성된 후 DNA의 특정 염기에 메틸화가 일어남. 주로 시토신(C) 염기에 메틸화 반응(15장의 불활성화 X염색체) • 전사가 덜 되는 세포에서 메틸화가 더 많이 일어남 • DNA의 메틸화와 히스톤 단백질의 탈아세틸화라는 이중 장치로 전사를 억제 • 15장의 유전자각인(genomicimprinting) 유전외적 유전(Epigenetic Inheritance) • 염색질의 변형 현상은 DNA 염기서열 그 자체의 변화를 수반하지는 않지만, 다음 세대로 전달됨. 이와 같이 염기서열 자체가 관여되지 않는 유전현상을 유전외적유전(epigeneticinheritance)라 함.
유전자 전사 개시 단계에서의 조절(Regulation of Transcription Initiation) 전형적인 진핵유전자의 구성(Organization of a Typical Eukaryotic Gene) • 그림 19.5: 전형적인 진핵세포 유전자와 그 발현을 조절하는 DNA 부위 • 조절요소(controlelement): 특정 단백질이 결합되어 유전자 전사 조절을 도와주는 DNA 부위 • 진핵 유전자는 다수의 조절요소와 연계
전사인자의 역할(The Roles of Transcription Factors) • 진핵세포의 RNA 중합효소II가 전사를 개시하기 위해 전사인자(transcriptionfactor)가 필요 • 17장에서 설명된 전사인자는 모든 단백질 코딩 유전자에 필요한 인자로서 일반 전사인자(general transcription factor, GTF)라 함 • 진핵세포에서 적당한 시간과 장소에서 특정 유전자의 전사가 대량으로 일어나기 위해서는 특수 전사인자(specific transcription factor)가 추가로 필요 • <Enhancers and Specific Transcription Factors> • 기부조절요소(proximal control element) 는 프로모터에 인저해 있고, 인핸서(enhancer)라 불리는 말초조절요소(distal control element)는 유전자의 위쪽 또는 아래쪽에 멀리 떨어져 위치(그림 19.5) • 특수 전사인자인 활성자(activator)는 인핸서에 붙어 전사를 촉진(그림19.6) • 수백 가지의 활성화 단백질는 2개의 부위를 가짐; (1) DNA 결합부위, (2) 활성화 부위 • 또 다른 특수 전사인자인 억제자(repressor)는 유전자 전사를 억제하는 역할 • <Combinatorial Control of Gene Activation> • 유전자의 수에 비해 조절요소의 수는 놀랄 정도로 적음 • 10여 가지의 짧은 뉴클레오티드 서열이 서로 다른 유전자의 조절요소에서 반복적으로 쓰임 • 인핸서는 평균 약 10개의 조절 부위를 가지며 각 부위는 하나 또는 둘의 특정 전사인자와 결합. 약 10개의 조절부위 염기서열로도 많은 수의 조합이 가능 • 그림19.7: 적은 수의 조절부위의 서로 다른 조합이 어떻게 전사조절을 하는가를 보여주는 예
조율되어 조절되는 유전자(Coordinately Controlled Genes) • 여러 진핵 세포의 유전체에 대한 최근 연구 결과, 동시에 발현되는 유전자들은 종종 같은 염색체 상에 집중되어 분포 • 특정 대사과정에 관여하는 효소 유전자들의 경우와 같이 조화되어 조절되는 유전자들이 종종 여러 염색체에 흩어져 존재하는데, 이런 경우에는 유전자의 조절부위 또는 그 조합이 동일하게 구성되어 있어 같이 조절됨. • 외부에서의 화학적 신호에 의해서 발현 조절이 유도(예, 스테로이드 호르몬이 세포 속의 특정 수용체와 결합하고, 이 복합체가 핵으로 들어가 전사인자로 작용)
전사 후 조절의 기작(Mechanisms of Post-Transcriptional Regulation) RNA 가공과정(RNA Processing) • 대체RNA스플라이싱(alternativeRNAsplicing)이 한 가지 예(그림 19.8) • 특정 세포에만 존재하는 조절 단백질이 전사체의 조절 부위에 결합하여 엑손-인트론 선택을 조절
mRNA 분해과정(mRNA Degradation) • 세포에서 단백질 합성 정도를 결정하는 주요 요인 중 하나는 세포질에서의 mRNA의 수명임 • 원핵세포 mRNA는 합성 후 수분 이내에 효소들에 의해 분해 • 다세포 진핵생물의 mRNA는 수 시간, 심지어는 수 주일까지 존속 • 효모에서의 연구에 따르면 mRNA 분해과정은 다중 (A) 꼬리가 짧아지는 것에서부터 시작 • mRNA 안정성을 결정하는 염기서열은 종종 3’-UTR에서 발견 • 최근 새로운 조절 기작으로 microRNA(miRNA)가 발견; miRNA는 긴 RNA 전구체로부터 만들어지는데, 이 전구체는 이중가닥의 머리핀 구조로 접혀 있고 다이서(dicer)라는 효소에 의해 잘려짐(그림 19.9). 이중 가닥 중 한 가닥은 분해가 되고 다른 가닥이 miRNA로서 커다란 단백질 복합체에 결합하여 상보적인 서열을 갖는 mRNA와 결합하게 됨. 이 결과 mRNA가 분해되기도 하고 번역이 저해됨 • RNA 분자에 의한 발현조절 현상은 이중가닥 RNA(dsRNA)(실제로는 DNA)를세포 속에 주입했을 때 동일한 서열을 갖는 유전자의 발현이 저해되는 것으로부터 알려짐(이를 RNA 방해(RNA interference, RNAi)라 하며 이는 작은 크기의 방해성 RNA(small interfering RNA, siRNA)에의한 것임이 밝혀짐)
번역의 시작(Initiation of Translation) • mRNA의 번역과정도 조절의 대상이 되는데 주로 개시단계가 조절의 표적(그림 17.17 참조) • mRNA의 5’-UTR에 있는 특정 서열 또는 구조에 특정 조절 단백질이 결합하여 리보솜의 결합을 방해함으로서 번역 개시가 저해될 수 있음 • 한 세포의 모든 mRNA의 번역을 동시에 조절할 수도 있음; 이러한 ‘글로벌’ 조절에는 번역 개시에 필요한 단백질에 대한 활성화 또는 불활성화가 관여 단백질 가공과정의 분해(Protein Processing and Degradation) • 이미 17장에서 설명 • 어떤 단백질이 세포 속에서 얼마 동안 역할을 할 것인가는 선택적인 분해작용에 의해 엄격하게 조절; 이를 위해 세포는 주로 유비퀴틴이라고 하는 작은 단백질을 대상 단백질에 부착시켜 표지하고, 프로테아좀(proteasome)이라고 하는 거대 단백질 복합체가 유비퀴틴으로 표지된 단백질을 인식하여 선택적으로 분해(그림 19.10)
개념 19.3: 암은 세포주기에 변화를 일으킬 수 있는 유전적 변화에 의해 발생할 수 있다 • 12장에서 암은 세포가 성장 조절 기작을 벗어나게 되어 발생되는 질병이라고 배웠음 암과 연관된 유전자 유형(Types of Genes Associated with Cancer) • 세포주기가진행되는 동안 세포의 성장과 분열을 조절하는 유전자로는 성장인자, 그 수용체 그리고 신호전달과정에 관여하는 유전자들이 있음. 이들 유전자 중 어느 것이라도 체세포에서 돌연변이가 일어나면 암이 될 수 있음 • 발암 바이러스 (tumor virus)가 인간을 포함한 여러 종에서 여러 종류 발견되었음(표 18.1 참조). 예를 들어, 파필로마바이러스(papilloma virus)는 cervix(자궁경부)암과 연관
발암유전자와 원발암유전자(Oncogenes and Proto-Oncogenes) • 암을 유발하는 발암유전자(oncogene). 이러한 유전자와 유사한 정상적인 유전자를 원발암유전자(proto-oncogene)이라 함. 이들 유전자는 주로 세포 성장과 분열을 촉진하는 단백질을 암호화 • 원발암유전자를 발암유전자로 바꾸는 변이는 크게 세 가지 부류; (1) DNA가 유전체 상에서 옮겨짐, (2) 유전자의 중복, (3) 유전자 그 차제 혹은 유전자 조절부위에서 점돌연변이의 발생(그림 19.11) 종양억제유전자(Tumor-Suppressor Genes) • 세포는 세포분열을 촉진하는 단백질의 유전자뿐 아니라 세포분열을 억제하는 단백질의 유전자도 갖고 있음. 이러한 유전자를 종양억제유전자(tumor-suppressor gene)라 하며, 이 기능이 저하되면 암의 발병이 일어날 수 있음
그림 19.11 원발암유전자가 발암유전자로 바뀌는 유전적 변화
정상적 신호전달의 저해(Interference with Normal Cell-Signaling Pathways) • 많은발암유전자와 종양억제유전자는 신호전달 과정의 단백질 정보를 암호화 • Ras유전자(Rasgene)(대표적인원발암유전자)는 G 단백질의 일종으로 신호전달의 결과 세포주기를 촉진하는 단백질을 생성(그림 19.12a). • P53(대표적인 종양억제유전자)는 발현 단백질이 전사인자로서 세포주기 진행을 저해하는 단백질의 합성을 촉진함(그림 19.12b) • P53의 돌연변이나 Ras의 과활성은 지나친 세포 성장을 초래하고 암을 유발(그림 19.12c) • P53유전자는단백질의 크기가 53Kda에 해당되며, “유전체의 수호천사(guardian angel of the genome)”이라불리며, DNA 손상 등에 활성화가 되면서 다양한 유전자의 전사를 촉진함.
암 발생과정의 다단계 모형(The Multistep Model of Cancer Development) • 암이돌연변이의 축적에 의해 발생하기 때문에 오래 살수록 암에 걸릴 확률이 높음. • 그림 19.13: 직장암에 대한 연구 • 세포가 암세포로 되기 위해서는 DNA 수준에서 5~6개의 변화가 일어나야 함. • 바이러스는 사람 암 발병에 약 15% 정도의 역할을 하는 것으로 조사 암의 유전적 소인(Inherited Predisposition to Cancer) • 어떤 암은 유전적 성향을 가짐; 예를 들어, 직장암의 약 15%는 유전되는 변이에 기인, 유방암의 경우에도 5~10% 정도에서 유전적 소임을 보임.
개념 19.4: 진핵세포의 유전체는 유전자 이외에 많은 수의 비암호화 DNA 염기서열을 가진다 • 진핵 유전체의 상당 부분은 비암호화 DNA 염기서열로 되어 있음. • 이를 “쓰레기 DNA(junk DNA)”라고부르기도 하지만 맞지 않음. 세포 속에서 중요한 일을 한다는 것이 밝혀지고 있으며, 진화의 오랜 시간 동안 이들 비암호화 DNA 부분이 사라지지 않은 것에는 이유가 있음 유전체의 성분과 개체 복잡성과의 관계(The Relationship Between Genomic Composition and Organismal Complexity) • 약간의 예외를 제외하고 대체로 작은 크기에서 큰 크기의 유전체로 진화 • 사람은 원핵세포에 비해 500~1,500배의 염기쌍을 가지지만 유전자의 수는 5~15배에 불과함. • 원핵세포에는 아주 적은 부분만이 비암호화 DNA 부분이며 주로 프로모터 등과 같은 조절부위임. • 사람의 유전체 분석이 완료된 결과, 유전체의 약 98.5%가 비암호화 DNA임(그림 19.14). • 유전자에 연관된 조절부위와 인트론 등은 약 24%이며 나머지는 유전자와 유전자 사이에 존재. 주로 반복서열 DNA(repetitive DNA)이며, 이들의 약 ¾는 전위인자(transposable element) 또는 그와 연관된 서열들임.
전위인자와 그와 연관된 서열(Transposable Elements and Related Sequences) • 모든 유전체는 여기저기 옮겨 다닐 수 있는 DNA 부위가 있음. • 미국의 매클린톡(McClintock)의 옥수수 연구에서 비롯(그림 19.15) 트랜스포존과 레트로트랜스포존의 이동(Movement of Transposons and Retrotransposons) • 진핵세포의 전위인자는 두 종류; DNA를 중간 매개로 유전체 사이를 옮기는 트랜스포존(transposon)과 RNA 중간물을 매개로 옮겨 다니는 레트로트랜스포존(retrotransposon) (그림 19.16)
그림 19.15 전위인자가 옥수수 알의 색에 미치는 효과
전위인자와 관련된 서열(Sequences Related to Transposable Elements) • 전위인자와 그 관련 서열이 포유류 유전체에서 차지하는 비율은 25~50%에 달하며 양서류와 일부 식물에서는 그보다도 더 높음. 단순서열 DNA를 포함하는 기타 반복 DNA(Other Repetitive DNA, Including Simple Sequence DNA) • 전위인자와관련이 없는 반복서열들은 아마도 DNA 복제 과정 등에서 실수로 도입되었을 것으로 추정. 인간 유전체의 약 15%를 차지하며 그 중 1/3은 크기가 큰 중복 DNA 부분으로 길이가 10,000~30,000 bp에 이름 • 단순서열 DNA(simple sequence)는 짧은 서열이 반복되어 연결된 형태. 반복단위가 수백 개씩 반복되어 있기도 함. 단순서열 DNA는 인간 유전체의 약 3%를 차지. 이를 위성 DNA(satelliteDNA)라고 하기도 함.
유전자와 다유전자군(Genes and Multigene Families) • 단백질과 구조 RNA를 암호화하고 있는 유전자는 불과 1.5%에 불과하며 그 유전자들에 있는 인트론 등을 포함하는 조절부위를 다 합쳤을 때 인간 유전체의 약 25%가 됨. • 다유전자군(multigenefamily): 동일 유전자나 유사한 유전자가 여러 개 있는 경우 • 히스톤 단백질을 제외하면 대부분 동일 유전자 다유전자군은 RNA를 암호화하고 있는 유전자들임 • 그림 19.17a: rRNA를 암호화하고 있는 유전자군 • 유사 유전자들의 다유전자군의 전형적인 예는 글로빈 단백질들을 암호화하고 있는 유전자군(그림 19.17b) • 위유전자(유사유전자; pseudogene): 유전자와 유사한 서열을 갖지만 실제로 발현되지 않는 서열
개념 19.5: DNA의 중복, 재배열 변이 등이 유전체의 진화에 기여할 수 있다 • 유전체 변화의 기본은 돌연변이이며 이는 유전체 진화의 기본이 됨. 유전체 크기의 확대 등 염색체의 복제(Duplication of Chromosome Sets) • 감수분열의실수가 염색체의 수를 증가시켜 다배수체(polyploidy)가 될 수 있음(24장 참고) DNA 조각의 중복과 다양화(Duplication and Divergence of DNA Segments) • 그림 19.18: 전위인자는 정확한 DNA 서열이 병렬되지 않은 상황에서도 교차 부위를 만들 수 있음. • DNA 복제 과정에서 실수가 있을 수 있는데, 주형이 되는 DNA를 두 번 읽거나 건너뛰는 실수가 생기면 DNA의 일부가 결실되거나 중복됨
그림 19.18 불균형적인 교차에 의해 생기는 유전자 중복
관련된 기능을 가지는 유전자의 진화: 글로빈 유전자의 예(Evolution of Genes with Related Functions: The Human Globin Genes) • 고대 단일 유전자가 4.5~5억 년 전에 복제되어 알파 글로빈과 베타 글로빈 유전자의 원형유전자로 나누어졌다고 생각됨(그림 19.19). 글로빈 수퍼유전자군(globin superfamily) • 인간의글로빈 단백질들 간의 유사성 비교(표 19.1) 새로운 기능을 갖는 유전자의 진화(Evolution of Genes with Novel Functions) • 교과서 참조. 리소자임(lysozyme) 유전자가 변화되어 알파 락트알부민(alpha-lactalbumin)으로 진화했다는 설명
그림 19.19 인간의 알파 글로빈과 베타 글로빈 유전자군의 진화
유전자 부분의 재배열: 엑손 복제와 엑손 셔플링(Rearrangements of Parts of Genes: Exon Duplication and Exon Shuffling) • 그림 19.20: TPA 단백질 유전자는 엑손 이동과 복제라는 기작으로 만들어진 것으로 추정 전위인자가 유전체 진화에 기여한 점(How Transposable Elements Contribute to Genome Evolution) • 전위인자가현재의 유전체에 많은 부분을 차지할 만큼 남아 있다는사실은 유전체 구성에 있어서 일정한 기여를 했을 것임을 짐작
그림 19.20 엑손 셔플링에 의한 새로운 유전자의 진화