Download
1 / 29
300 likes | 530 Views
6. Alternative splicing controls sex determination in Drosophila. Sexually dimorphic characteristics. Alternative splicing controls sex determination in Drosophila. Polyadenylation of the 3’ end of the nascent transcript is required for stability. Polyadenylation occurs in two phases:

E N D
Alternative splicing controls sex determination in Drosophila Sexually dimorphic characteristics
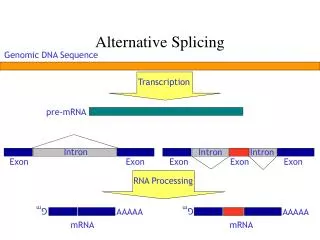
Alternative splicing controls sex determination in Drosophila
Polyadenylation of the 3’ end of the nascent transcript is required for stability • Polyadenylation occurs in two phases: • A slow phase mediated by Poly A Polymerase (PAP)during which approximately12 A residues are addedon to the cleaved 3’ end. • This structure is recognised byPoly A Binding Protein II (PABPII-to distinguish it from the cytoplasmic PABP),which catalyses therapid addition of ~200 A residues.
Splicing variants or isoforms provide added diversity: Alternative Splicing
How Is the Information Content in DNA Transcribed to Produce RNA? The genetic code: specifies which amino acids will be used to build a protein Codon: a sequence of three bases. Each codon specifies a particular amino acid. Start codon: AUG—initiation signal for translation (translational start site) Stop codons: stops translation and polypeptide is released (translational termination signal)
Figure 12.8 Transfer RNA anticodon
Pre-tRNAs undergo critical processing Pre-tRNAs are transcribed by RNA Pol III and thereafter are processed to yield mature tRNA • 1) the 5’ end sequence is removed • 2) a short intron is removed • 3) CCA is added on to the 3’ end • 4) extensive modification of internal bases
Since rRNA accounts for approximately 80% of the total RNA in the cell, it has to be transcribed extensively to be able to satisfy the continuous demand Pre-rRNA transcription units are arranged in repetitive clusters
the arrangement 5'-3' 18S - 5.8S - 28S is invariant within and between species Eukaryotic rRNA is transcribed in units comprising the 18S, 5.8S and 28S rRNAs
thepre-rRNA transcript is extensively processed in the nucleolus. Not only is it cleaved into its components, but alsonumerous bases are chemically modified. all thepre-rRNA processing is executed through small nucleolar RNAs, which are part of snoRNPs. unlike the other rRNAs encoded by the single pre-rRNA molecule,5S rRNAis transcribed outside the nucleolus and arrives there by diffusion andis not further processed Precursor rRNA is extensively processed
the overall stem-loop structure of ribosomal RNA is highly conservedamong the thousands of species investigated to date. (molecule shown is 16S bacterial rRNA, similar to 18S eukaryotic rRNA) rRNA folds into conserved stem-loop structures
rRNA is an integral part of the protein synthesizing machine, the ribosome
Assembly of the pre-initiation complex Protein synthesis can be negatively regulated by phosphorylation of eIF2.
two types of tRNAs exist for the methionine codon tRNAiMet is exclusively used for initiation of a polypeptide chain tRNAMet is exclusively used for elongation of a polypeptide chain The tRNAiMet is needed to start synthesis of a polypeptide chain the methionine in bacterialtRNAi is modified by addition of a formyl group both are charged by the same aminoacyl tRNA synthetase
Assembly of the pre-initiation complex Protein synthesis can be negatively regulated by phosphorylation of eIF2.
Because thedimeric guanyltransferase bindsto thephosphorylatedC-terminal domain (CTD) ofRNA polymerase II only mRNA transcripts are capped! therefore,only class II (mRNA) transcripts are efficiently translated! A 5' cap is required for the efficient translational initiation of eukaryotic mRNAs
A cap-binding protein is required for efficient translational initiation • Sonenberg et al. purified a protein that bound the 7mGDP cap specifically using an affinity chromatography approach • Addition of the purified protein to capped mRNAs increased the translational efficiency in vitro, but did not affect uncapped transcripts. X
eIF4 is instrumental in recruiting an mRNA to the preinitiation complex • next,a protein complex named eIF4 binds the pre-initiation complex as well as the cap of an mRNA. • An associated helicase activity removes any secondary structure in the 5’ region of the transcript...
The initiation complex uses ATP energy and the helicase activity of eIF4A to scan the mRNA until the initiation codon is found (usually the first AUG). eIF4E-Cap eIF4G-binds eIF3 eIF4A-RNA helicase (ATP) ACC G
eIF4 and mRNA are recruited to form an initiation complex Activity of the cap-binding subunit eIF4E can also be regulated.
the ribosome has to recognize the start codon of the ORF the ribosome start site for protein synthesis is directed toward the AUG initiator codon by the Shine-Dalgarno box, a 5-8bp sequence that can base pair with the 16S rRNA moleculein the small ribosomal subunit Bacterial translation initiation requires the Shine-Dalgarno box
the pairing of the Shine-Dalgarno box with the 16S rRNAresults in a match of the initiation AUG codon with the initiator tRNA (tRNAi) The Shine-Dalgarno box positions the AUG initiation codon for pairing with the tRNAi in mammalian cells a relatively conserved sequence ACCAUGG recognises the start site (AUG)- KOZAK sequence (after Marilyn KOZAK)
Recruitment of the large ribosomal subunit completes the initiation phase. Commitment to translation (can no longer dissociate)