Download
1 / 35
370 likes | 636 Views
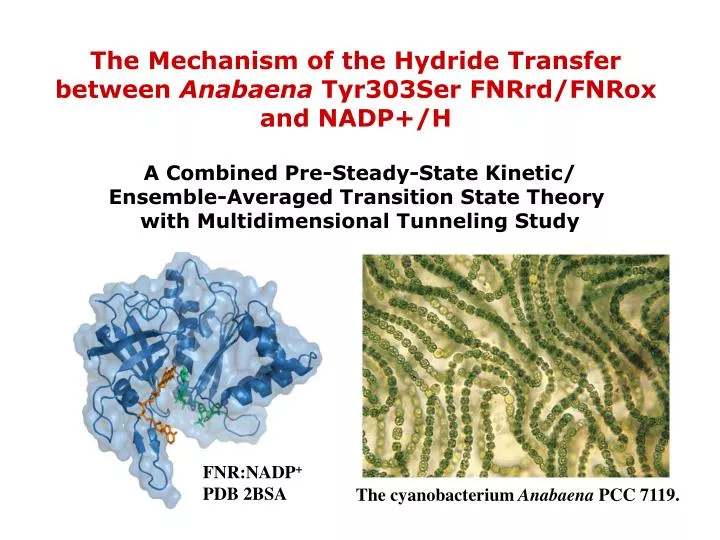
The Mechanism of the Hydride Transfer between Anabaena Tyr303Ser FNRrd/FNRox and NADP+/H A Combined Pre-Steady-State Kinetic/ Ensemble-Averaged Transition State Theory with Multidimensional Tunneling Study. FNR:NADP + PDB 2BSA. The cyanobacterium Anabaena PCC 7119.

E N D
The Mechanism of the Hydride Transfer between Anabaena Tyr303Ser FNRrd/FNRox and NADP+/H A Combined Pre-Steady-State Kinetic/ Ensemble-Averaged Transition State Theory with Multidimensional Tunneling Study FNR:NADP+ PDB 2BSA The cyanobacterium Anabaena PCC 7119.
José M. Lluch, Mireia Garcia-Viloca and Àngels Gonzàlez-Lafont Departament de Química and Institut de Biotecnologia i de Biomedicina, Universitat Autònoma de Barcelona, 08193 Bellaterra (Barcelona), Spain. Isaias Lans, José Ramón Peregrina and Milagros Medina Departamento de Bioquímica y Biología Molecular y Celular, and Institute of Biocomputation and Physics of Complex Systems (BIFI), Universidad de Zaragoza, E-50009, Zaragoza, Spain. Darmstadt September 05 - 09, 2010.
Cytosol Phycobilisoms CO2 Fixation N2 Metabolism ATPase PSI Cytb6f PSII FNR Fdox+ PSIrd Fdrd + PSIox 2 Fdrd + NADP+ + H+ 2 Fdox + NADPH The cyanobacterium Anabaena PCC 7119 and its photosynthetic chain
1e- 2e- + H+ Fe2S2 NADP+ Electron Transfer Chains in Biological Systems FAD
3 4 2 1 two Fdrd molecules + H+ NADP+ FNR 2 Fdrd+ NADP+ + H+ 2 Fdox + NADPH
NADP+ Y303 FAD FAD NADP+ A B Hydride Transfer Mechanism Reduction of NADP+ to NADPH is proposed to take place by hydride transfer from the N5 position of the flavin ring of FNR to the C4 position of the nicotinamide ring. Tyr303seems not to be involved in the hydride transfer process but prevents the direct interaction between the flavin ring of FNR and the nicotinamide ring of NADP+. Tyr303 side-chain must be displaced from its position during hydride transfer. FNR(1GJR) Y303S FNR(2BSA) N5-C4N 3.4 Å N5-C4N 7.78 Å N10-N1N 4.6 Å Rings 30º
By means oftwo complementary studies: Theoretical study based on EA-VTST/MT Experimental stopped-flow pre-steady-state kinetic study OBJECTIVES A better understanding of how FNR enzyme works, that is, of how the hydride transfer process that FNR catalyzes takes place in Tyr303Ser FNR To shed light on the role of C-terminal Tyr303 in WT FNR Lans, I. et al. J. Phys. Chem. B2010, 114, 3368 – 3379
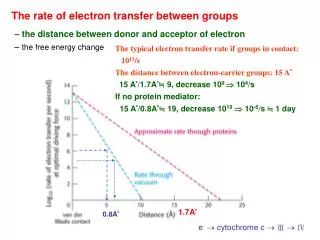
EXPERIMENTAL RESULTS T=279 K 285 s-1 270 s-1 ’ The equilibrium mixture is displaced towards NADPH production, consistent with the physiological main role of the enzyme. 190 s-1 Tyr303 substitution by a Ser practically deactivates the capacity of the enzyme to reduce NADP+ BUT: Tyr303 in the WT must be displaced for the hydride transfer to take place !!
NADP+ Anabaena Tyr303Ser FNR:NADP+ complex 2BSA BUT:The structural disposition in the mutant between the flavin and the nicotinamide rings seems favorable to produce the hydride transfer.
OBJECTIVES of the theoretical approach Provide very subtle details of the mechanism that could be unavailable with experimental techniques. By means of a fully microscopical simulation of the hydride transfer in the complete solvated FNR:NADP+ system. Calculate the macroscopic rate constants and kinetic isotope effects of the hydride transfer in FNR that could be compared with the experimental data. By means of Ensemble-Averaged Transition State Theory with Multidimensional Tunneling.
Gas-phase model hydride-transfer reaction: stacking lumiflavin (FADH-/FAD) 1-methylnicotinamide (NADP+/NADPH) AM1 fails to describe the dispersion energycontribution to stacking favoring instead intermolecular hydrogen-bonding. MPWB1K/6-31+G(d,p) accounts better for stacking interactions than B3LYP (6 kcal/mol of energy difference in complexation energies). At the MPWB1K/6-31+G(d,p)level lumiflavin and 1-methylnicotinamide are in a vertical stacked configuration.
COMPUTATIONAL DETAILS Model of the biological system Initial Cartesian coordinates PDB file 2BSA. Protonation states: PROPKA. H atoms added with HBUILD in CHARMM. System neutralized with 7 Na+ Cubic box of water molecules. Total number of atoms:41896 atoms (4800 protein atoms) Equilibration of the solvated protein/cofactor/coenzyme system CHARMM22 force field+TIP3P NPT molecular dynamics (MD) simulations with periodic boundary conditions (PBC) at 279 K and 1 atm. Particle Mesh Ewald method. SOFTWARE: CHARMM35 1-D QM/MM Potential Energy Profile ABNR algorithm Mobile part: all atoms within a sphere of 20 Å. Distinguished coordinate z.
H- QM Definition of the QM region (atoms in orange) used for the QM/MM calculations Total number of QM atoms: 58 Frontier GHO atoms: 2 Total number of MM atoms : 41838
RD RA SP FNRrd-NADP+ FNRox-NADPH 1D-POTENTIAL ENERGY SURFACE: AM1(QM)/CHARMM22-TIP3P(MM) . The hydride transfer HT-1 in the complete solvated enzymatic system results in: An endoergic process (7.8 kcal/mol) with a potential energy barrier of 36.7 kcal/mol. The flavin and the nicotinamide rings are set out in a roughly parallel configuration.
Dual level single-point energy correction , High Level Low level .
COMPUTATIONAL DETAILS CHARMMRATE =CHARMM + POLYRATE MPWB1K//AM1(QM)/CHARMM22-TIP3P(MM) NPT molecular dynamics (MD) simulations with periodic boundary conditions (PBC) at 279 K and 1 atm. Particle Mesh Ewald method. Potential of mean force (PMF) and classical free energy curve with the umbrella sampling technique along the reaction coordinate z and the weighted histogram analysis method (WHAM). .
35.8 kcal/mol 34.9 kcal/mol = 0.04 Ả = -0.07 Ả CLASSICAL POTENTIAL OF MEAN FORCE The introduction of the MPWB1K energy correction only changes significantly the endergonicity of the reaction. 14.9 kcal/mol 3.6 kcal/mol The location of the transition state moves slightly towards products. The effect of thermal and entropic contributions is small.
33.3 kcal/mol 14.7 kcal/mol QUASICLASSICAL POTENTIAL OF MEAN FORCE Quantized-vibration correction It is worth noting that the vibrational contribution lowers the relative free energy of the transition state around 2.5 kcal/mol.
EA-VTST/MT rate constants: activation free energy barriers Classical mechanical activationfree energy profile Quasiclassical activation free energy profile (kcal/mol) Quantal vibrational corrections clearly reduce the free energy barriers. The quassiclassical free energy barrier for the physiological reaction HT-1 is almost twice as big as the corresponding value for the reverse reaction HT.
Structural analysis along the reaction coordinate H TS N5 R N10 C4N N1N 4.6 Ả N10 –N1N The N5 – hydride – C4N angle has to approach to 180º, and the hydride donor and acceptor atoms have to come closer. Deformation of the contact ion pair FNRrd-NADP+ and partial loss of π stacking interaction 3.4 Ả 2.75 Ả N5 – C4N An energy penalty that increases the free energy cost of the transition state
Z2 Z1 R SP P Average net transmission coefficient γ For each variational transition state configuration i at z*QC: Static-Secondary Zone approximation (Frozen Bath) • AM1/CHARMM22 SP location • AM1/CHARMM22MEP • Dual-level MEP energy correction • (interpolation using ISPE). • 4) Quassiclassical transmission factor Γi • 5) Semiclassical transmission coefficient κi
Vi(s) for configuration i. i(T) = exp{-DGi} DDGi Potential of mean force tunneling I(T) Reaction Coordinate z Quassiclasical Transmission Factor Γ
Semiclassical transmission coefficient P quantum = = P classical Veff Pcl(E) 1 Veff,* Pqu(E) 1/2 E E VAG s< s> s Pqu(E) Action integral along the tunneling path
= 1 = 0 s Tunneling path RBC • MEP • Assuming vibrational adiabacity of F-1 modes orthogonal to s RAB E • Zero-Curvature (ZCT) tunneling
Reaction-path curvature Coupling of vibrations k to the reaction coordinate s “corner cutting” Small-curvature (SCT):
(kcal/mol) EA-VTST/MT rate constants:dynamic effects CHARMMRATE Average net transmission coefficient i ≡ configuration of VTSE Quassical transmission factor Semiclassical transmission coefficient: SCT
HT-1 35.04 31.87 0.998±0.003 335.70±142.79 334.82±142.44 2.2X10-10 28.64 HT 20.08 17.02 0.998±0.003 335.70±142.79 334.82±142.44 92.8 13.80 EA-VTST/MT rate constants: (s-1) Phenomenological free energy of activation (kcal/mol) The hydride transfer from Tyr303Ser FNRrd to NADP+ hardly occurs in agreement with our stopped-flow kinetic measurements. Conversely, the reverse reaction, HT, does happen, with a reaction rate constant of 92.8 s-1 in very good agreement with our stopped-flow kinetic measurements (kHT = 190 s-1).
H D A B Hydride and deuteride transfer in the Tyr303Ser FNR:NADP+ reactant complex: Kinetic Isotopic Effect in the non-photosynthetic direction Ln(kobs) = lnA - Ea/RT QM/MM H/D 10.73 kcal/mol FAD:NADPH (C4-H) FADH-:NADP+ (N5-H) 10.68 kcal/mol TUNNEL HYDRIDE TRANSFER Vibration-driven tunneling
Conclusions • The physiological hydride transfer from Tyr303Ser FNRrd to NADP+ is not possible. The reverse reaction, the hydride transfer from NADPH to Tyr303Ser FNRox,does occur. • The experimental and theoretical reverse reaction rate constants are in very good accordance. • At the reactant region the N5-C4N distance might be compatible with the hydride transfer, but the N5-hydride-C4N angle is very far from collinearity, and therefore, the hydride shift is quite inefficient. In going from the reactant region to the transition-state region the N5-hydride-C4N angle approaches 180º to make the hydride transfer easier, and the hydride donor and acceptor atoms come closer. • Since the width of the hydride transfer reaction path is small, the hydride transfer involves an important degree of quantum mechanical tunneling. • The H/D KIE is essentially temperature-independent. • All those geometric deformations, including a partial loss of the π stacking interaction, involve a large free energy penalty to reach the transition state. The difference between the direct and the reverse reaction rates comes from the important positive reaction free energy (15 kcal/mol). Such a difference must be due to an important stabilization of the close contact ionic pair FADH-:NADP+ versus the situation corresponding to the product, where, after the hydride transfer, the two moieties are neutral.
WT FNR Possible role of Tyr303 in WT FNR? Tyr303Ser FNR Any factor able to distort the formation of that close contact ionic pair would destabilize the reactant, so increasing the rate constant of the direct reaction. Very interestingly, the affinity for the coenzyme in the WT FNR (Kd = 5.7 μM),is much lower than in the mutant Tyr303Ser FNR (Kd < 0.01 μM), this fact being consistent with the feasibility of the direct hydride transfer corresponding to the physiological main role of the enzyme. Could this lower affinity in the WT FNR be attributed to the presence of Tyr303 distorting in some way the formation of a close contact ionic pair as stable as in the Tyr303Ser mutant, with the corresponding thermodynamic and kinetic consequences?
-2011 Chair Persons: Manuel Yáñez Otilia Mó Co-chair: Saulo Vázquez Contact: watoc2011@uam.es
Catalytic mechanism of hydride transfer. Role of transient charge transfer interactions. • charge-transfer complex • An electron-donor–electron-acceptor complex, characterized by electronic transition(s) to an excited state in which there is a partial transfer of electronic charge from the donor to the acceptor moiety. • charge-transfer (CT) transition • An electronic transition in which a large fraction of an electronic charge is transferred from one region of a molecular entity, the electron donor, to another, the electron acceptor (intramolecular CT) or from one molecular entity to another (intermolecular CT). Mulliken, R. S. (1952), J. Am. Chem. Soc. 74, 811 l
FNRhq FNRsq FNRqn Fdrd (Fldhq) Fdox (Fldsq) NADPH NADP+ electrons PSI Transient Interactions in the photosynthetic electron transfer from PSI to NADP+
Corrección vibracional H Corrección vibracional D E(kcal/mol) z z Ajuste a una curva gaussiana Corrección Vibracional