Download
1 / 63
640 likes | 817 Views
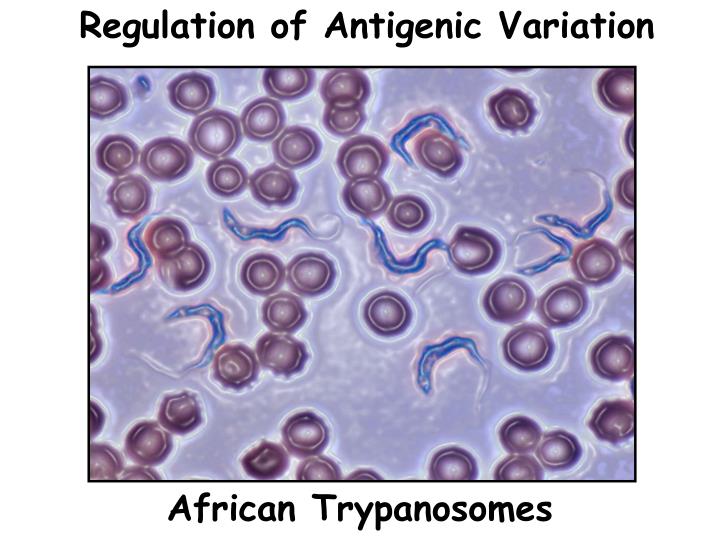
Regulation of Antigenic Variation. African Trypanosomes. VSG. switch. Antigenic Variation. stochastic. in vitro = in vivo. Immune. destruction. by host. Proliferation. Variant Surface Glycoprotein. 1000 diff variants. Only one expressed. on the surface. 20 nm thick. VSG. switch.

E N D
Regulation of Antigenic Variation African Trypanosomes
VSG switch Antigenic Variation stochastic in vitro = in vivo Immune destruction by host Proliferation Variant Surface Glycoprotein 1000 diff variants Only one expressed on the surface 20 nm thick
VSG switch Regulation of Antigenic Variation Immune destruction by host Proliferation Stochastic No influenced by immune system 1,000 VSG genes VSG VSG genes Telomere VSG expression site
VSG switch Duplicative transposition Regulation of Antigenic Variation Immune destruction by host Proliferation 1,000 VSG genes VSG VSG genes Telomere VSG expression site
VSG switch Duplicative transposition Regulation of Antigenic Variation Immune destruction by host Proliferation 1,000 VSG genes VSG VSG genes Telomere VSG expression site
Antigenic variation systems use subtelomeric localization Telomeric repeat Subtelomere First chromosome- internal gene TTAGGGTTAGGGTTAGGG….. Protect chromosomes: Degradation End-end fusions Benefits of subtelomeric localization: Recognition by DNA repair Reversible gene silencing Loss of info by replication Telomere position effect (TPE) Allow rapid switching and exclusive expression Ectopic recombination Increase gene family diversification Rapid gene switching Why telomeric location ???
Subtelomeric localization of VSG gene in T. brucei VSG X (19) VSG Benefits of subtelomeric localization: Need to regulate both Reversible gene silencing Telomere position effect (TPE) Allow rapid VSG switching and exclusive expression Ectopic recombination Increase gene family diversification Rapid VSG gene switching
Antigenic Variation 7 6 2 1 ESAGs VSG VSG genes 50 bp 70 bp telomeric ~ 1,000 Repetitive DNA sequences
Antigenic Variation 7 6 2 1 (1) VSG VSG genes active X 6 7 2 1 (19) VSG inactive VSG switching 1. Recombination mechanisms duplicative transposition telomeric exchange 2. In situ switch
Antigenic Variation X 7 6 2 1 (19) VSG inactive VSG genes 6 7 2 1 (1) active VSG VSG switching 1. Recombination mechanisms duplicative transposition telomeric exchange 2. In situ switch How to follow switch events ??
Chromosome mapping of switch events Telomere exchange Duplicative transposition WT Null WT Null WT Null 221 1.3 221 1.3 221 1.12 SLOT 221 ES 221 ES Probe 221 1.3 1.12 Laura Cliffe
Why multiple Expression Sites? 6 4 7 2 7 6 2 1 (1) 50 bp 70 bp ESAGs VSG genes X 6 7 2 1 (19) (~200) 177 bp 1. Recombination Dominant mechanism duplicative transposition telomeric exchange 2. In situ switch
Transferrin receptor hetero-dimer Expressed on surface: flagella pocket binds iron
Transferrin receptor in the flagellar pocket Tf-R ESAGs VSG 7 6 5 4 8 3 2 1 VSG surface coat flagellum mitochondrion Tf-R VSG nucleus flagellar pocket kinetoplast Membrane of the flagellar pocket Allows the uptake of iron
Why Multiple Expression Sites ? 6 4 2 7 Cow Each ESAG 6 and 7 in ESs are Slightly different X 7 6 4 2 Dog Resulting in receptors with differing affinities for transferrin from different hosts X 7 6 4 2 Sheep X 7 6 4 2 Human Trypanosomes growing in Cow serum-----------put into dog serum ????
Why Multiple Expression Sites ? ESAG 6 and 7 X 6 4 2 7 Cow Hetero-dimer transferrin receptor 7 6 4 2 Transferrin is an iron containing molecule essential for growth Dog X 7 6 4 2 Each ESAG 6 and 7 are slightly different Sheep X 7 6 4 2 Differences reflected in differing affinities for transferrin Human Experiment: Trypanosomes growing in Cow serum-----------put into dog serum ???? Select for trypanosomes that have switched
Why Multiple Expression sites ? X 6 4 2 7 Cow 7 6 4 2 Dog X 7 6 4 2 Sheep X 7 6 4 2 Human Allowed host range expansion What is the consequence of choice during infection ?
Host range of Trypanosoma brucei Buffalo Spotted hyena Waterbuck Wild dog Buffalo Coke’s Hartebeest Spotted hyena Wild dog Coke’s Hartebeest Waterbuck Eland Lion Giraffe Zebra Reedbuck Giraffe Zebra Reedbuck Eland Lion Warthog Bushbuck Impala Hippo Cattle Warthog Bushbuck Impala Hippo Cattle
Host range of Trypanosoma brucei Buffalo Spotted hyena Waterbuck Wild dog Buffalo Coke’s Hartebeest Spotted hyena Wild dog Coke’s Hartebeest Waterbuck Eland Lion Giraffe Zebra Reedbuck Giraffe Zebra Reedbuck Eland Lion Warthog Bushbuck Impala Hippo Cattle Warthog Bushbuck Impala Hippo Cattle
Multiple ESs leads to multiple Mechanisms 6 4 2 7 (1) 50 bp 70 bp ESAGs VSG genes ~ 1,000 X 6 4 7 2 (19) (>100) 177 bp 1. Recombination Dominant mechanism duplicative transposition telomeric exchange 2. In situ switch Early infection
Mechanisms of VSG Switching Monomorphic Pleomorphic 667, 927 427 Strain Lab adapted Lifecycle Experimental line of choice (?) 10-2 - 10-5 10-6 - 10-7 Switch rate slow rapid Recombination Recombination Switch Mechanism In situ switch In situ switch
Regulation of Antigenic Variation (1) VSG VSG genes active ~ 1,000 X (19) VSG inactive Why is only one expression site active ? ESB localization How are the inactive ESs stably repressed ? Lack of Pol I ?
Regulation of Antigenic Variation (1) VSG VSG genes active ~ 1,000 X (19) VSG inactive Why is only one expression site active ? ESB localization How are the inactive ESs stably repressed ? Lack of Pol I ? Active repression mechanisms chromatin structure ? Regulation of telomeric VSG DNA recombination ? DNA modification chromatin structure
Histone methytransferase Dot1 H3 lysine 76
DNA methylation and histone modification MBP MBP 5MeC DNA binding proteins * * Recruit Histone methylase 3HC 5-methylcytidine
Histone Code Dot1 K79
Chromatin structure X 7 6 2 1 (19) VSG inactive 6 7 2 1 (1) active VSG ISWI (ATPase/helicase) ‘molecular motor’ Slight de-repression of ES transcription
Chromatin structure X 7 6 2 1 (19) VSG inactive 6 7 2 1 (1) active VSG ISWI (ATPase/helicase) Slight de-repression of ES transcription
Dot1: Histone H3K76 methylase X 7 6 2 1 (18) VSG inactive 6 7 2 1 (1) active VSG X 7 6 2 1 VSG inactive Dot1 KO (disruption of telomeric silencing) Slow rate of silencing previously active ES
Dot1: Histone H3K76 methylase X 7 6 2 1 (18) VSG inactive 6 7 2 1 active VSG Slow silencing 7 6 2 1 active VSG Dot1 KO (disruption of telomeric silencing) Slow rate of silencing previously active ES
How are the inactive ESs stably repressed? X X X PvuII PstI PvuII (1) VSG active 1984 PstI PvuII PvuII X (19) inactive Blocked Restriction Sites Within Silent ESs DNA Modification?
The Novel Base J J-DNA Base J Glucose Thymine 1993 J.H. Gommers-Ampt
Consequence of glycosylated DNA ???? J J J J J J J J J J J J J J J J J 70 bp VSG 50 bp J J J J J J J J J J J J J J J J J X J J J J J J J J J J J J J J J J J X
Knock-Out J-Synthesis Pathway in T. brucei 1993 structure (1) VSG active X 1999 (19) localization X 2007 X Base J function Loss of VSG gene regulation ??
Regulation of J-Biosynthesis HOMeUra DNA synthesis Thymidine- hydroxylase Glucosyl- transferase (TH) (GT) HOMedU b-D-glucosyl-HOMedU dT BS PC - + HMU Why do procyclics lack J ? 100 ng Lack thymine hydroxylase (?) 50 25 DNA titration 12 6 3 1.5 Anti-J dot blot
Regulation of J-Biosynthesis HOMeUra DNA synthesis Thymidine- hydroxylase Glucosyl- transferase (TH) (GT) HOMedU b-D-glucosyl-HOMedU dT BS PC - + HMU Why do procyclics lack J ? 100 ng Lack thymine hydroxylase (?) 50 25 DNA titration 12 JBP1 J-DNA binding TH 6 JBP2 TH SWI2/SNF2 3 1.5 Thymine-hydroxylase motif Anti-J dot blot
JBP1 Binds J-DNA and Stimulates J Synthesis JBP1 KO ACCCTAACCCTAACCCTAAC TGGGATTGGGATTGGGATTG Tel-T * Tel-OH Tel-J Tel-T + JBP1 JBP -/- WT -OH ACCCTAACCCTAACCCTAAC TGGGATTGGGATTGGGATTG Tel-OH Bound * DNA titration Glc -O- ACCCTAACCCTAACCCTAAC TGGGATTGGGATTGGGATTG Free Tel-J * Kd = 90 nM J-DNA binding JBP1 H D H R L L L 189 191 239 255 406 411 431 How does it bind J-DNA ??
JBP2 Induces De-Novo J-Synthesis JBP1 JBP2 120 BS PC 85 WT J1KO WT J1 J2 Tubulin Tubulin Phleo GFP JBP array array Anti-GFP JBP1 Stimulate J synthesis J-DNA binding JBP2 SWI2/SNF2 Stimulates de-novo J ATP-dependent 34% identity 47% similarity DNA helicase
JBP2 Induces De-Novo J-Synthesis JBP1 JBP2 120 85 Tubulin Tubulin Phleo GFP JBP array array Anti-GFP BS PC PC WT J1KO WT J1 J2 J2 J2/J1 JBP1 J-DNA binding Propagates J synthesis TH JBP2 SWI2/SNF2 TH Stimulates de-novo J Thymine-hydroxylase motif AlkB-like hydroxylase domain
Thymidine hydroxylase domain of JBP2 / JBP1 ? HO JBP1 JBP2 Thymine Hydroxymethyluracil O2 CO2 + + 2-oxoglutarate Succinate HCHO OH Fe2+ AlkB Thymine 3-methylthymine
Iron and 2-Oxoglutarate Hydroxylase Family Conserved structural fold: eight -strands Iron and Oxoglutarate binding H1-X-D-Xn-H2-Xn-R motif Essential residues Strategy: Mutate JBP1 and JB2 J-DNA binding TH TH SWI2/SNF2 H-X-D-Xn-H-Xn-R H189A D191A H239A Stimulate J-biosynthesis? R255A V258A Zhong Yu et. al., NAR 2007 : Laura Cliffe et. al., NAR 2009
JBP2 Stimulates HOMedU Formation in E. coli Anti-HOMedU IP assay E. coli expression E. coli DNA - + IPTG 7 8 190 7 6 120 6 5 5 4 85 CPM x 104 CPM x 104 4 3 3 60 2 2 50 1 1 0 1 mM 5 mM + HMU -JBP2 IPTG Fe2+ DNA oligos *pHOMedU OH JBP2 12 ..ATTGCT.. ..ATTGCT.. *pA 10 8 CPM x 104 *pT 6 4 *pG 2 Unmod HOMedU Anti-HOMedU IP Robbie Southern
Regulation of J-Synthesis by Two Thymidine Hydroxylases O2 CO2 + + Succinate 2-oxoglutarate Glc O HO Fe2+ GT JBP1 JBP2 J Thymine Hydroxymethyluracil Why are two thymidine hydroxylases needed ??
Deletion of JBP2 and JBP1 Ablates J-Biosynthesis X T T-OH T-O-Glc TH GT +JBP2/ JBP1 J2 J1 ddKO JBP1 dKO JBP2 dKO WT BF +HMU WT PC Glucosyl-transferase still present JBP2 and JBP1 rescue JBP2/JBP1: Thymidine hydroxylases -J dot blot Rudo Kieft and Laura Cliffe
Lack of Base J and Regulation of VSG Expression 221 6 7 2009 (1) Base J function 6 7 X J Null cell line (19) 6 7 X 1. De-regulation of expression sites 2. Increased VSG switching rate 3. Repeat stability
J null cells 7 6 Active 7 6 Active? 7 6 Active? Does J play a role in expression site silencing ? WT cells VSG VSG 7 6 Active J J J J J J J J X VSG VSG 7 6 Silent X J J J J J J J J VSG VSG 7 6 Silent ~90% 6/7 mRNA from 221ES RT-PCR analysis VSG 7 6 Cross-reactive primers
J plays no role in Transcriptional silencing J null cells VSG 7 6 Active X % transcripts from 221 ES VSG 7 6 Silent X VSG 7 6 Silent J null WT Laura Cliffe