Download

1 / 22
230 likes | 576 Views
Mekanisme Molekuler Induksi Tumor Crown Gall oleh Agrobacterium tumefaciens. I. PENDAHULUAN UMUM. TIK: Mahasiswa memahami sifat-sifat Agrobacterium tumefaciens. Mahasiswa memahami penyakit Crown Gall pada tanaman yang diinduksi oleh Agrobacterium tumefaciens.

E N D
Mekanisme Molekuler Induksi Tumor Crown Gall oleh Agrobacterium tumefaciens
I. PENDAHULUAN UMUM TIK: • Mahasiswa memahami sifat-sifat Agrobacterium tumefaciens. • Mahasiswa memahami penyakit Crown Gall pada tanaman yang diinduksi oleh Agrobacterium tumefaciens. • Mahasiswa memahami proses-proses yang terlibat dalam induksi tumor Crown Gall. Waktu : 2 x 50menit
Sub Pokok Bahasan: 1.1 Pendahuluan 1.1.1 Penyakit Crown Gall 1.1.2 Sifat-sifat Agrobacterium tumefaciens 1.1.3 Induksi Crown Gall oleh Agrobacterium 1.2 Proses-proses yang Terlibat dalam Induksi Tumor Crown Gall 1.2.1 Penempelan Agrobacterium pada Sel Tanaman 1.2.2 Induksi Gen-vir 1.2.3 Proses dan Transfer T-DNA 1.2.4 Penggabungan T-Strand ke dalam Genom Tanaman
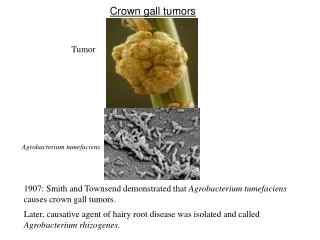
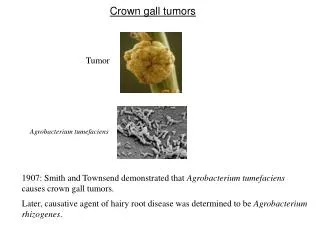
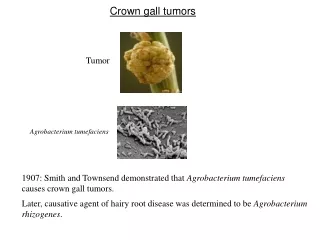
1.1 PENDAHULUAN 1.1.1 Penyakit Crown Gall • Tumor Crown Gall adalah jaringan tanaman yang pertumbuhannya tidak terdiferensiasi akibat adanya interaksi antara tanaman-tanaman yang rentan dengan strain virulen Agrobacterium tumefaciens (Gb. 1).
Karakterisasi molekuler dari induksi Crown gall ini menunjukkan bahwa Agrobacterium bisa dipakai untuk mengantarkan materi genetik kedalam tanaman dan hal ini menjadi ketertarikan baru dalam studi Agrobacteriumtumifaciens. • Sekarang ini, sistem transfer DNA dari Agrobacterium ke tanaman dimanfaatkan secara meluas untuk penelitian biologi molekuler dan rekayasa genetika pada tanaman.
1.1.2 Sifat-sifat Agrobacterium tumefaciens • Agen penyebab (causative agent) penyakit Crown gall, yaitu Agrobacterium tumifaciens, digolongkan kedalam famili Rhizobiaceae yang dibagi seperti ditunjukkan pada Gb.2. • Sebagian besar genus Agrobacterium menyebabkan tumor pada tanaman dikotil. Spesies Agrobacterium tergolong bakteri gram-negatif yang merupakan bakteri aerob dan mampu hidup baik sebagai saprofit maupun parasit.
Agrobacterium berbentuk batang, berukuran 0.6-1.0 µm sampai 1.5-3.0 µm, dalam bentuk tunggal atau berpasangan. • Agrobacterium merupakan bakteri yang mudah bergerak (motile) dan memiliki 1-6 flagela peritrichous serta merupakan bakteri tak berspora. Suhu optimal pertumbuhan bakteri ini adalah 25-28◦C. • Kumpulan bakteri ini biasanya berbentuk cembung, bulat, lembut, dan tak berpigmen. (46)
A. tumafaciens mengandung sebuah plasmid besar yang disebut Ti-plasmid, yang bertanggung jawab pada proses onkogenesitas A. tumefaciens. Namun, Ti-plasmid sendiri tidak mampu menyebabkan terjadinya transformasi pada tanaman. • Gen-gen pada kromosom A. tumifaciens juga memiliki fungsi lain yang berguna untuk onkogenesitas in planta (57, 65). • Ti-plasmid hidup secara stabil dalam tubuh bakteri meskipun ukurannya besar (200-900 kb) (33). Struktur dari Ti-plasmid ditunjukan oleh Gb. 3. • Wilayah virulen (vir) pada Ti-plasmid, menyediakan sebagian besar produk trans-acting untuk transit T-DNA.
1.1.3 Induksi Crown gall oleh A. tumefaciens • Peristiwa pembentukan tumor Crown gall yang diakibatkan oleh Agrobacterium digambarkan pada Gb. 4. • A. tumefaciens menyerang tanaman pada bagian yang luka. Pengirim pesan kimia (chemical messengers), biasanya senyawa fenolik seperti acetosyringone, yang berasal dari sel tanaman yang luka, menginduksi transkripsi gen virulen Ti plasmid-borne di dalam bakteri (5, 72). • Produk-produk gen vir mempengaruhi transfer T-DNA ke sel tanaman. T-DNA mengkode enzim auksin, sitokinin, biosintesis; fitohormon ini menggangu keseimbangan hormon pada tanaman inang, sehingga terbentuklah tumor.
1.2 PROSES-PROSES YANG TERLIBAT DALAM INDUKSI TUMOR CROWN GALL Tumorigenesis dari Crown gall merupakanproses yang terdiri dari beberapa tahap: • penempelan Agrobacterium pada sel tanaman • proses dan transfer T-DNA ke sel tanaman • penggabungan dan pengekspresian gen-gen T-DNA di dalam genom tanaman.
1.2.1Penempelan Agrobacterium pada Sel Tanaman • Tahap pertama dalam induksi tumor adalah pengikatan Agrobacterium pada sel tanaman. Peristiwa awal ini dimediasi oleh gen-gen yang terletak pada kromosom bakteri. • Gen-gen kromosom, chvA, chvB, dan exoC, diperlukan dalam sintesis dan penyaluran β-1,2-glucancyclic yang terlibat dalam pengikatan sel tanaman. • Protein chvB terlibat dalam biosintesis glucan (67), sementara chvA diperlukan untuk memindahkan glucan dari sitoplasma ke periplasma dan ruang ekstraseluler.
1.2.2 Induksi Gen-vir • Daerah vir kira-kira 30 kb dan tersusun dalam 7-8 komplementasi kelompok:virA, virB, virC, virD, virE, virF, virG, dan virH (sebelumnya pinF) (73, 86). • Gb. 5 menunjukan susunan genetik wilayah vir dari Ti-plasmid tipe oktopin dan nopalin. Perbedaan wilayah vir antara Ti-plasmid tipe oktopin dan nopalin adalah; virF dan virH (pinF) tidak terdapat pada Ti-plasmid tipe nopalin sementara tipe oktopin mengandung kedua gen tersebut.
Dua penginduksi potensial, acetosyringone dan hydroxy-acetosyringone, diisolasi dari sel akar tembakau oleh Stachel et al (72). • pH medium memiliki pengaruh yang kuat pada induksi vir; induksi akan optimal dalam rentang pH 5.0-5.4 dan tidak akan terjadi induksi pada pH 6.3 atau diatasnya (74). • Regulasi gen vir juga dimediasi oleh gen-gen yang terletak pada kromosom Agrobacterium. Gen virulen kromosomal yang dinamai chvD, chvE, ros, miaA, chvG, dan chvI telah dilaporkan terlibat dalam virulensi dengan cara mempengaruhi ekspresi gen-gen vir pada Ti-plasmid (85, 86, 89,36,28, 18, 10,51).
1.2.3 Proses dan Transfer T-DNA • Proses dan transfer dari T-DNA dimediasi oleh produk-produk yang dikode oleh gen-gen pada wilayah vir (vir-region). Setelah induksi vir gen, produksi transfer tingkat lanjut dimulai dengan pembentukan T-strand, yaitu duplikat dari single stranded (ss) T-DNA (74). • Banyak produk gen bertanggung-jawab dalam proses ini. Produk dari loci virC dan VirD terlibat dalam pembentukan dan pemrosesan T-strand. VirD1 dan VirD2 bersama-sama mengenali susunan pembatas 25 bp dan memproduksi sebuah belahan endonukleotik ss pada strand bagian bawah dari masing-masing pembatas (25, 27). Sayatan ini digunakan sebagai tempat inisiasi dan terminasi dalam memproduksi T-stand.
Setelah penyayatan, VirD2 tetap terhubung kuat dengan ujung 5’ dari T-strand (35). Pengikatan VirD2 pada ujung 5’ memberi karakter polar pada T-strand yang menjamin bahwa pada langkah berikutnya, ujung 5’ merupakan leading end. • Produksi T-strand diperkirakan merupakan hasil dari perpindahan strand bagian bawah dari T-DNA di antara sayatan tersebut. • Studi terbaru mengungkapkan bahwa langkah awal produksi T-strand secara evolusioner terkait dengan sistem bakteri yang memproduksi ssDNA, seperti salah satunya selama proses konjugasi (60).
Dua protein lainya, VirC1 dan VirC2 kemungkinan berinteraksi dengan VirD1 dan VirD2 selama penyayatan berlangsung. Lokus virC, yang mengkode dua produk yaitu VirC1 (23 kDa) dan VirC2 (26 kDa), digunakan untuk meningkatkan proses penyayatan batas dari T-DNA (81, 82, 89). • T-strand harus bergerak melalui membran dan ruang seluler sebelum sampai pada inti sel tanaman dan tetap menjaga integritasnya selama proses ini. Oleh karena itu, adalah sebuah hipotesis bahwa T-strand seolah-olah bergerak seperti sebuah kompleks protein-ssDNA.
VirE2 merupakan sebuah protein yang memiliki ikatan asam nukleat ss yang dapat diinduksi (inducible ss nucleic acid-binding protein), yang dikode oleh lokus virE yang terikat tanpa kespesifikan susunan. • Sejumlah besar VirE2 (60 kDa) berada dalam sel dan terikat kuat serta saling bekerja sama, yang berarti bahwa sebuah T-strand akan terlapisi secara keseluruhan (Gb. 7). • Satu molekul VirE2 diperkirakan melapisi sekitar 30 nukleotida, oleh karena itu 20 kb T-strand akan membutuhkan 600 molekul VirE2 (52).
Akibatnya, degradasi T-strand oleh nukleasi akan dicegah dengan ikatan VirE2 dan bahkan, ikatan in vitro virE2 membuat ssDNA resisten terhadap degradasi nuckleolitik. • T-strand bersama-sama VirD2 dan virE2 membentuk T-kompleks. T-kompleks merupakan kompleks nukleoprotein yang sangat besar. Di beberapa strain Agrobacterium, T-strand panjangnya mencapai 20 kb (65). T-strand ini akan mengikat sampai sekitar 600 molekul virE2 dan memiliki panjang terhitung 3600 nm, yaitu 60 kali ketebalan membran inti. Termasuk sebuah VirD2, T-kompleks memiliki massa 50x106 D (70). • Kemudian, T-komplek harus keluar dari sel bakteri, melewati membran dalam dan luar dan juga dinding sel bakteri (Gb. 7).
1.2.4 Integrasi T-strand ke Genom Tanaman • Integrasi T-strand ke dalam sebuah kromosom inang merupakan tahapan akhir dalam proses transfer T-DNA (Gb. 7). • Integrasi T-strand membutuhkan pengambilan inti (nuclear uptake).Protein yang berukuran lebih besar dari kira-kira 40 kDa membutuhkan NLS (nuclear localization signal) yang memediasi pengambilan inti (34).
Analisis susunan VirD2 mengungkapkan bahwa pada wilayah ini ditemukan susunan yang homolog dengan NLS tipe bipartite (34). Jika sebuah NLS bipartite yang tidak berpasangan dari VirD2 kurang mampu memediasi pengambilan inti pada T-kompleks yang sangat besar, makadiperlukan tambahan NLSs yang dapat dihasilkan dari VirE2, komponen protein yang paling berlimpah dari T-kompleks. Analisis sekuensing menunjukkan dua NLSs bipartite yang potensial dalam VirE2 (12). • Jadi, VirE2 menyediakan NLSs di sepanjang T-kompleks. NLSs ganda pada VirE2 kemungkinan bermanfaat secara fungsional untuk impor inti T-kompleks yang tak terputus/terganggu, seperti contohnya, untuk menjaga sisi sitoplasma dan nukleoplasma secara simultandari pori-pori inti yang terbuka.
Mekanika integrasi T-strand ke dalam genom tanaman sebagian besar belum diketahui. Disarankan bahwa ikatan VirD2 dengan ujung 5’ pada T-strand bergabung dengan sayatan pada DNA tanaman. DNA tanaman akan lebih terbuka untuk membentuk sebuah celah, dan ujung 3’ dari T-strand kemungkinan berpasangan dengan wilayah lain dari DNA tanaman. Reaksi-reaksi ini akan menghasilkan T-strand ke dalam satu strand dari DNA tanaman. Strain yang terpilin kemudian akan menimbulkan sebuah sayatan pada strand yang berlawanan dari DNA tanaman. • Reparasi celah dan sintesis DNA menggunakan T-strand sebagai cetakan, menghasilkan produk integrasi akhir. Namun, belum diketahui apapun tentang enzimologi proses integrasinya. Tidak ada faktor-faktor tanaman yang terlibat dalam proses ini. • VirD2 dan VirE2 dianggap mengkatalisasi integrasi T-strand kedalam genom tanaman; namun, tidak ada data yang mendukung hipotesis ini.
DAFTAR GAMBAR • Gb. 1. Induksi Crown Gall oleh A.tumefaciens (strain A208) pada stem Kalanchoe daigremontiana • Gb. 2. Hubungan anggota-anggota famili Rhizobiaceae (46) • Gb. 3.Organisasi genetic Ti plasmid tipe octopine. • Loci vir dan T-DNA dijelaskan pada penjelasan. Loci-loci yang lain adalah sebagai berikut: tra, transfer konjugasi, occ, katabolisme octopine; arc, katabolisme arginine; agc, katabolisme agropine; inc, ketidakcocokan; ori, asal pengulangan (86) • Gb. 4. Gambaran peristiwa-peristiwa induksi Crown Gall oleh A.tumefaciens. Penjelasan lebih lengkap pada teks. • Gb. 5. Organisasi genetic wilayah vir pada Ti plasmid octopine (A) dan Ti plasmid nopaline (B). Panah-panah di atas menunjukan unit transkripsi. Dua gen (pinF dan virF) yang ada pada Ti plasmid octopine tidak ada pada Ti plasmid nopaline (85) • Gb. 6. Sisterm regulator gen-gen vir pada Agrobacterium. Lihat teks untuk penjelasan lebih lengkap. OM, membrane luar; IM, membrane dalam. • Gb. 7. Tahapan dasar dalam transformasi sel-sel tanaman oleh A.tumefaciens (91)