Download
1 / 26
410 likes | 1.81k Views
Mus musculus. Homo sapiens. Pisum sativum. Amoeba dubia. Xenopus laevis. Escherichia coli. Lilium longiflorium. Caenorhabditis elegans. Protopterus aethiopicus. Mycoplasma pneumoniae. Saccharomyces cerevisiae. Drosophila melanogaster. The C-value paradox.

E N D
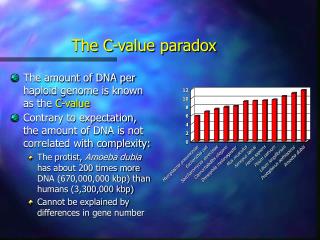
Mus musculus Homo sapiens Pisum sativum Amoeba dubia Xenopus laevis Escherichia coli Lilium longiflorium Caenorhabditis elegans Protopterus aethiopicus Mycoplasma pneumoniae Saccharomyces cerevisiae Drosophila melanogaster The C-value paradox • The amount of DNA per haploid genome is known as the C-value • Contrary to expectation, the amount of DNA is not correlated with complexity: • The protist, Amoeba dubia has about 200 times more DNA (670,000,000 kbp) than humans (3,300,000 kbp) • Cannot be explained by differences in gene number
The structure of genes • There are many forms of genes: • Those which produce a protein, a tRNA or an rRNA are referred to as structural genes • Those which control how and when genes are expressed are called regulatory genes • Some housekeeping genes need to be expressed in all tissues e.g. those involved in protein synthesis • Other, tissue-specific genes, are only expressed in a particular cell or tissue type e.g. the insulin gene is only expressed in the pancreatic β-cells • Whatever their function, all genes contain a coding region which specifies a polypeptide or an RNA molecule
Promoter region • TATA box • CAAT box (in mammals) • GC box (GGGCGGG) • Promoter region • Shine-Dalgarno box (AGGAGG) • Pribnow box (TATAAT) • -35 site (TTGACA) Polyadenylation signal AATAA Eukaryote Exon 1 Exon 2 Exon 3 Exon 4 5’ 3’ Intron 1 Intron 2 Intron 3 Initiation codon Initiation codon Stop codon Stop codon Prokaryote 5’ 3’ Structure of a typical gene - alcohol dehydrogenase (Adh)
Generating Genetic Variation • Gene Duplication: the globin family • A classic example of gene duplication and evolution • Globin molecules are involved in carrying oxygen in multicellular organisms • Ancestral globin gene (present in primitive animals) was duplicated ~500 mya. • Mutations accumulated in both genes to differentiate them - α and β present in all higher vertebrates • Further gene duplications produced alternative forms in mammals and in primates
Primates Mammals
Generating Genetic Variation • Gene Duplication • Almost every gene in the vertebrate genome exists in multiple copies • Gene duplication allows for new functions to arise without having to start from scratch • Studies suggest the early in vertebrate evolution the entire genome was duplicated at least twice
Generating Genetic Variation • Exon Duplication • Duplications are not limited to entire genes • Proteins are often collections of distinct amino acid domains that are encoded by individual exons in a gene • The separation of exons by introns facilitates the duplication of exons and individual gene evolution
Generating Genetic Variation • Exon Shuffling • The exons of genes can sometimes be thought of as individual useful units that can be mixed and matched through exon shuffling to generate new, useful combinations
Gene duplication in the Hox gene family • Homeotic genes control the development of body plan in animals • In both vertebrate Hox and invertebrate HOM genes, there is a highly conserved protein motif known as a homeobox • Mutations in Hox/HOM genes can drastically affect the organisation of body parts
Gene duplication in the Hox gene family • Although Hox/HOM genes are related, their organisation differs between organisms: • In vertebrates, there are multiple clusters of Hox genes: the mouse has four clusters, each located on a different chromosome and covering over 100 kb • HOM genes in Drosophila are found in two clusters, Antennipedia and Bithorax, on the same chromosome • In amphioxus – a class of marine invertebrates which are the closest relatives to the vertebrates – there is a single cluster of at least 10 Hox genes each of which is homologous to a different Hox gene in vertebrates: origin of vertebrates coincided with a series of gene duplications • Example of a dispersed gene family in vertebrates
Gene Duplications (four clusters) Amphioxus Hypothetical Common Ancestor Drosophila lab pb Dfd Scr Antp Ubx AbdA AbdB Gene duplication in the Hox gene family
Introns • Occur frequently within eukaryotic genomes and make up most of the length of very long genes • Number, size and organisation of introns varies: • Histones have no introns: chicken pro-a2-collagen gene has over fifty • SV40 virus contains an intron of 31 bp: human dystrophin gene has an intron of over 210,000 bp • Some introns have genes contained within them - the Adh gene in Drosophila is located within the intron of the outspread gene • Strong conservation of intron-exon boundaries - nearly always begin with GT and end with AG
Types of introns • Most introns in eukaryotes are spliceosomal introns (‘nuclear introns’) because they are spliced by a spliceosome of proteins and RNA • Some introns can splice without the aid of proteins (“self-splicing introns”): • One class - group I introns - are sometimes mobile because they encode proteins such as DNA endonucleases. They are found in mitochondrial and chloroplast genomes, rRNAs of some eukaryotes and in T4 bacteriophage • Group II introns are found in organelles and their bacterial ancestors and contain reverse transcriptase-like sequences • Group III introns are found in a few protists and are similar to group II introns with the central portion removed
The evolution of introns • There are two competing hypotheses for the evolution of spliceosomal introns: • The introns-early hypothesis, proposed by Walter Gilbert, suggests that introns mark the boundaries between ancient genes which encoded distinct proteins. • Throughout evolution these once-independent proteins have been put together in new combinations to produce more complex proteins by exon shuffling • An alternative hypothesis (introns-late) suggests that introns only invaded eukaryote genomes fairly recently
The evolution of introns (continued) • A crucial prediction of the introns-early hypothesis is that spliceosomal introns delineate structural or functional units within proteins: • Introns are found in the same places in all known globin genes, including myoglobin and plant leghaemoglobins • More frequently, however, introns do not appear to separate functionally distinct parts of proteins • Other problem with introns-early hypothesis is absence from Archaea and Bacteria: • Massive intron loss has been postulated but does not explain why they are found in nuclear copies of organelle genes but not in the genes of the organelles or their precursors • Exon shuffling has probably been a factor in later eukaryotes