Download

1 / 42
470 likes | 1.91k Views
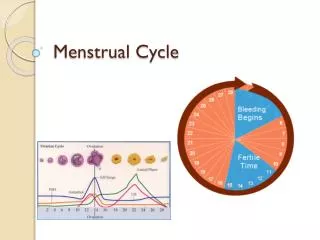
Menstruation & ovulation. PHYSIOLOGY OF MENSTRUAL CYCLE. The normal menstrual cycle is divided into: 1. The ovarian cycle. 2. The uterine cycle. THE OVARIAN CYCLE.

E N D
PHYSIOLOGY OF MENSTRUAL CYCLE • The normal menstrual cycle is divided into: • 1. The ovarian cycle. • 2. The uterine cycle.
THE OVARIAN CYCLE • The changes that occur in the ovary during each cycle can be divided into three stages: (1) The follicular phase (day 1 to 13), (2) Ovulatory phase (day 13 to 15) and (3) The luteal phase (day 15 to 28).
The Follicular phase: • During the follicular phase, certain number of follicles start to grow and some of them will pass from the stage of primordial follicle to the stage of preantral follicle, and usually few of them can pass to the antral follicle stage of which one only succeeds to continue through the pre-ovulatory follicle stage. In each of these stages, there are many cellular, histological, hormonal and functional changes that occur with interaction between the gonadotropins and the ovarian steroid hormones.
The primordial follicle: • During the intrauterine life, the ovarian differentiation starts in-between the 6th and the 8th week by proliferation and multiplication of the germ cells to reach a huge number (6-7 millions) around the 20th week. The germ cells stop mitotic division and start meiosis under the effect of secretions of the mesonephric tubules-derived cells "Rete cells" called meiosis inducing substance. Passing through the early stages of meiosis, the oocytes will be arrested at the prophase of meiosis and become surrounded by a layer of perivascular cells that are both mesenchymal and epithelial in origin to form the primordial follicles. Failure of achieving this coating by the granulosa cells would lead germ cell to complete meiosis and die with marked reduction of the germ cell population. This process requires the presence of two X chromosomes and so, in 45XO chromosomal pattern of Turner's syndrome, gonadal dysgenesis occurs.
The primordial follicle is composed of oocyte arrested in the diplotene stage of meiotic prophase, surrounded by a single layer of pregranulosa cells (germinal epithelial) resting on a basement membrane separating it from an outer less organized matrix of pretheca cells (mesenchymal cells). The rest of mesenchymal cells not utilized in primordial follicle formation is noted in the interstices between follicles, forming the primitive ovarian stroma. • The process of follicular maturation is described as a Continuum. It means that (The initial stages of follicular growth occur during all physiological conditions without any external stimulation). Each follicle seems to have a genetic code for the timing at which it may resume growth. The number of follicles that resumes growth at any point of time is dependent on the size of the residual pool of follicles in both ovaries and if the residual pool is changed dramatically at any time, the remaining follicles will rescheduled their readiness to resume growth according to the new situation.
The first visible sign that the follicle resumes growth is that the granulosa cells become cubical rather than squamous in shape. Mitosis appears in the granulosa cells, the oocyte expands and starts to secrete the zona pellucida (ZP). The overall diameter of the primordial follicle is about 50 and the oocyte is about 20 . Gap junctions develop between surrounding granulosa cells and the oocyte to establish a metabolic and electric linkage that will persist at ovulation. Not all granulosa cells are receptor +ve, so activation signals, protein kinase and cyclic AMP can pass from receptor +ve granulosa cells to receptor -ve cells. • Once the primordial follicle resumes growth, the process is irreversible, so if the situation is favorable, it would passes to the next stage, otherwise, atresia is the definitive fate. So the number of follicles in the ovaries is reduced dramatically as time passes. The general pattern is initial growth followed by atresia which is interrupted at the beginning of the cycle during menses or even during the last very few days of the previous cycle where a group of emerging follicles is exposed, responds to the rising peptide hormones stimulation ( withdrawal of the -ve feedback as the CL degenerates) and is propelled to further growth.
The maximum number ( 6-7 millions ) is present at the 20th week of intrauterine life, of which 2 millions are present at birth. No similar rate of depletion of the germ cell mass is seen again, and there is evidence that the major mechanism for this loss is by elimination through the surface of the ovary into the peritoneal cavity. At the onset of puberty, the germ cell mass has been reduced to 300,000. During the next 30-45 years of reproductive life, these units will be depleted further to a point at menopause where follicles are almost completely depleted. As 300-500 follicles would grow enough to reach the ovulation phase, for each of them close to 1,000 will pursue abortive growth periods of variable length.
The Preantral Follicle: • Once growth is initiated, the follicle passes to the preantral stage where the oocyte enlarges to reach a size of 80 ( same size till ovulation, limited by the ZP ). The granulosa cells undergo multilayer proliferation and the theca cells begins to organize from the surrounding stroma with the development of its own newly formed blood vessels. The overall diameter of the follicle at this stage is about 200. • The growth in this stage is dependent on the gonadotropin stimulation. LH acts mainly on the theca cells to start steroidogenesis and the production of androgens mainly testosterone and androstenedione. These androgens may pass directly to the blood stream, but also some of it will back-diffuse toward the granulosa cell layer where it is aromatized by the granulosa cells under the effect of FSH. FSH not only stimulates the conversion of androgens to estrogen but also propels follicles to the antral stage by stimulating granulosa cell mitosis. Together, FSH and estrogen increase the FSH receptors content of the follicle with a resulting snowballing effect on follicle growth and differentiation.
Androgen plays a delicate role to determine the fate of the follicle. The presence of moderate amounts of androgen in the follicle not only acts as a substrate but also via androgen specific receptor- stimulates its aromatization to estrogen while excessive androgen would act as anti-estrogen to inhibit aromatization leading to androgen accumulation converted to 5-reduced form which can not be converted to estrogen , so, the follicle becomes androgenic and ultimately atretic.
Antral Follicle: • As the follicle continues its growth under the combined effect of FSH and estrogen, follicular fluid accumulates in the intercellular spaces to form Call-Exner bodies and coalescing together to form the characteristic follicular cavity "The Antrum" . The antrum provides a specific hormonal environment for the oocyte and the avascular granulosa cell layer which is different from that of the serum and the other follicles. The oocyte now bulges in the antrum being surrounded by few layers of granulosa cell called cumulus oophorus which attaches the oocyte to the rest of the granulosa cell. The theca cell layer is differentiated to well organized highly vascular theca interna and less organized less vascular theca externa.
The antral follicle is characterized by the highest level of granulosa cell proliferation, highest rate of FSH activity through huge number of FSH receptors, and so the greatest ability to convert androgen to estrogen making the antral follicle an estrogen-dominant follicle with accumulation of FSH and estrogen in the antrum. This is accompanied by a high degree of theca cell layer vascularization which allows a preferential delivery of FSH and LH to this follicle. The successful conversion to an estrogen-dominant follicle marks the selection of the follicle destined to ovulate "the dominant follicle", a process by which, except in rare exception, only one follicle ovulates each cycle. The natural selection of the dominant follicle is evident by day 7 of the cycle, although the process of selection started at an earlier stage when one of the primordial follicles has good synchronization between its timed resumption of growth and the elevated level of FSH seen during the last few days of the previous cycle.
This emerging dominant follicle produces about 95% of the entire ovarian secretion. Within the antral follicle, estrogen interacts with FSH to help maturation of the dominant follicle, while estrogen released in the circulation has a -ve feedback effect on the production and release of FSH from the anterior pituitary leading to a gradual drop in circulating FSH level. The drop in FSH level would reduce aromatase activity in less mature, less vascularized follicles and reduce granulosa cell proliferation and activity promoting accumulation of androgen thereby inducing irreversible atretic changes. The dominant follicle is immune from the drop in FSH as it has its own reservoir of FSH in the antrum with the highest granulosa cell mass and the highest degree of FSH receptors. So a wave of atresia among smaller follicles is seen in parallel to the rise of circulating estrogen produced from the dominant follicle.
Preovulatory Follicle: • At this stage the follicle attains a big size, it measures 20 mm or more. The granulosa cells enlarge and attain lipid inclusion, while the theca cells are highly vascularized with even more luteinization. • The oocyte resumes the nuclear component of meiosis I. It passes from the late stages of the prophase to the metaphase, anaphase and finally to the telephase of meiosis I. It is at this stage when the haploid number of chromosomes is produced as one pair of the chromosomes together with a little cytoplasm forms the first polar body that lies free inside the zona pellucida, while the second pair with the rest of the cytoplasm forms the secondary oocyte. Meiosis I is completed during this stage and the secondary oocyte enter meiosis II immediately before ovulation. Meiosis II is only completed at the entry of the sperm at the time of fertilization.
It is at this stage when the hypothalamic-pituitary ovarian interaction is at the most critical point with great synchrony and harmony: • Estrogen, mainly from the dominant follicle, reaches a high level and it should reaches a level of 200 pg/ml or more in the plasma to be maintained for at least 50 hours in order to produce the positive feedback on the pituitary which induces the LH surge at midcycle. The peak value of estrogen is reached 24-36 hours before ovulation while FSH declines to a nadir level. • LH starts to rise 32-38 hours before ovulation and reaches a peak value 10-12 hours before it. Acting upon its own receptor which were induced by the combined effect of FSH and estrogen, LH starts luteinization of the granulosa cells resulting in the production of progesteron within the microenvironment of the preovulatory follicle 24-48 hours before ovulation. Gonadotropins released at midcycle are characterized with high bioactivity and longer half life with increased content of sialic acid. • Progesterone in very small amounts facilitates the positive feedback response of the pituitary to threshold level of estrogen with enhanced pituitary response to GnRH. In addition to this, progesterone is responsible for the FSH surge at midcycle.
FSH has a midcyclic surge as LH but of much lesser amplitude which serve to ensure that a full complement of LH receptors is in place in the granulosa cells. • Androgens are also produced from the ovary at this stage. The theca component of the other follicles which fail to achieve full maturity, return to their origin as a component of the ovarian stroma. They retain their ability to respond to LH stimuli with the production of androgen in the ovarian stroma. The plasma androstenedione rises by 15% and testosterone by 20% at midcycle. This rise in androgens acts locally to assure and complete the wave of atresia in the non-dominant follicles and may has a systemic effect to increase lipido at midcycle.
Ovulation: • Ovulation or follicular rupture is not due to increased intra-follicular pressure. The escape of the ovum is preceded by separation of the oocyte-cumulus cell mass from the rest of the follicle to float freely in the antral fluid with massive increase in the amount of the fluid. The escape of the ovum is associated with degenerative changes of the collagen in the follicular wall which occurs just prior to ovulation. This is accompanied by completion of the oocyte maturation and luteinization. The midcyclic LH surge is the main stimulus to ovulation, but it should be synchronized with oocyte maturation and other morphological and functional changes within the dominant follicle. AS the main stimulus for the LH surge is the high estrogen level produced by the dominant follicle, it is the follicle itself that determines the proper timing of ovulation. In addition to LH, the FSH surge, estrogen and progesterone all play a role in the process of ovulation through manipulating the activity of several non steroid products in the follicle, this may include:
LH induced rise in cyclic-AMP overcomes the effect of oocyte maturation inhibitor (OMI) and luteinization inhibitor (LI) which are two non steroid products of the follicle present in the follicular fluid and serve to inhibit premature oocyte maturation and luteinization. • LH and progesterone stimulate proteolytic enzymes (collagenase and plasmin), resulting in the digestion of collagen in the follicular wall. This may be mediated via the production of prostaglandins E and F which may act to free lysosomal enzymes. • Prostaglandins may also stimulate the contraction of smooth muscles which is present in the ovary. • FSH is a good stimulator for the release of plasminogen activators which are responsible for the activation of plasminogen to plasmin. FSH stimulates the synthesis of hyaluronic acid whose accumulation is necessary in the separation of the oocyte-cumulus cell mass from the rest of the follicle.
The midcyclic increase in LH is short-living and it shortly ends with a decline in the level of LH. The exact mechanism is not yet fully known, but it may be due to exhaustion of the LH storage in the pituitary, -ve feedback of LH itself on the GnRH pulse generator, down regulation of GnRH receptors, or loss of the +ve feedback stimulus as estrogen level plunges as LH reaches its peak with a precipitous drop in circulating estrogen level.
Luteal phase: • After ovulation and escape of the oocyte with the cumulus cell mass to the peritoneal cavity, dramatic morphological changes occurs in the remaining part of the follicle within the next 2-3 days converting it to the corpus luteum. The name is derived from deposition of lutein, a yellow pigment of fat cells and egg yolk, inside the granulosa cells which become vacuolated and in the theca-lutein cells differentiated from the surrounding theca and stromal cells. • Another very important anatomical change is the penetration of the granulosa cells for the first time by blood capillaries from the underlying theca-lutein layer, with filling of the cavity with blood. Vascularization reaches its peak 8-9 days after ovulation. This change is very important to deliver LDL to the granulosa cells from which they extract cholesterol needed for steroidogenesis.
The cells of the corpus luteum which have been prepared during the follicular phase by FSH and estrogen and have a great number of LH receptors produces great amounts of both estrogen and progesteron under LH stimulation. Continuos LH stimulation at low level is needed to maintain the steroid production by the CL. Progesterone plasma level reaches its peak level 8 days after the LH surge. • Progesterone acts centrally through the hypothalamus to suppress gonadotropin production, and locally it suppress new follicular growth in the ovary containing the CL. This effect is mediated by depletion of estrogen receptors which is needed to support early follicular differentiation. So during the next cycle, usually the dominant follicle will be present in the contralateral ovary. • The duration of the luteal phase is consistently close to 14 days from the LH surge to menses. In the absence of pregnancy, the CL would decline within 9-11 days after ovulation with decline in the steroid production. Luteolysis is induced by the high level of estrogen reached in the second half of the luteal phase. This effect is mediated by estrogen stimulated production of prostaglandins within the ovary which uncouple the LH receptor complex from the adenylate cyclase enzyme across the lipid bilayer of the cell membrane. IB pregnancy occurs, HCG, produced from the chorion starts to rise 9 days after ovulation to rescue the CL from regression.
THE ENDOMETRIUM (UTERINE) CYCLE The histology of the adult endometrium: • The endometrium can be divided into an upper 2/3 functionalis layer and lower 1/3 basalis layer on the basis of morphology and function. • The purpose of functional layer is to prepare for implantation of blastocyst. It is the site for (1) proliferation, (2) secretion and (3) menstruation. • The purpose of basalis layer is to provide the regenerative endometrium following menstrual loss of functionalis.
The histologic changes are based on two parts: • Endometrial glands. • Stroma.
Proliferative phase: • In the proliferative phase, tissue components (glands including endothelial cells, stromal cells) demonstrate proliferation, which peaks on days 8-10 of the cycle, corresponding to peak estradiol levels in the circulation and maximal estrogen receptor concentration in the endometrium. This proliferation is marked by increased mitotic activity and increased nuclear DNA and cytoplasmic RNA synthesis, that is most intense in the functionalis layer in the upper two-thirds of the uterus, the usual site of blastocyst implantation. • At the beginning, the endometrium is relatively thin (1-2 mm). The initially straight narrow and short endometrial glands changes to longer tortuous structure. The organization changes: low columnar pattern early proliferative (5th day of the cycle) to a pseudo-stratified before ovulation (12th day of the cycle); the stroma is a dense compact layer throughout this time. Vascular structure is infrequently seen.
Secretory phase: • The corpus luteum produces large quantities of progesterone which induces secretory changes in the glands and swelling of stromal cells. There is a rich blood supply and the capillaries become sinusoidal with little intervening stroma. • The first histologic sign that ovulation has occurred is the appearance of subnuclear intracytoplasmic glycogen vacuoles in the glandular epithelium on cycle days 17-18. Giant mitochondria and the "nucleolar channel system" appear in the gland cells. Individual components of the tissue continue to display growth, but confinement in a fixed structure leads to progressive tortuosity of glands and intensified coiling of the spiral vessels. These structural alterations are soon followed by active secretion of glycoproteins and peptides into the endometrial cavity. Transudation of plasma also contributes to the endometrial secretions. The peak secretory level is reached 7 days after the midcycle gonadotropin surge, coinciding with the time of blastocyst implantation.
Of note the endometrial height is fixed at roughly its preovulatory extent (5-6 mm) despite continued availability of estrogen. Epithelial proliferation ceases 3 days after ovulation. This restraint or inhibition is believed to be induced by progesterone. • Implantation phase (Late Secretory); ie 7th - 13th post ovulation (21th - 27th of cycle) whereby the distended tortuous secretory glands have been most prominent with little intervening stoma. The time of implantation is in days 21-22 of the cycle. The predominant morphologic feature is edema of endometrial stroma secondary to the estrogen- and progesterone-mediated increase in prostaglandin production. By day 13-14, post-ovulatory: the endometrium is divided into 3 distinct zones: 1/4 unchanged basalis, 1/2 stratum spongiosum, 1/4 stratum compactum superficial layer.
Menstrual phase: • The menstrual endometrium is a relatively thin but dense tissue. It is composed of the stable, nonfunctioning basalis component and a variable, but small, amount of residual stratum spongiosum.The menstrual endometrium is a transitional state bridging the more dramatic proliferative and exfoliative phases of the cycle. Its density implies that the shortness of height is not entirely due to desquamation. Collapse of the supporting matrix also contributes significantly to the shallowness. Nevertheless, as much as two-thirds of the functioning endometrium is lost during menstruation. The more rapid the tissue loss, the shorter the duration of flow. Delayed or incomplete shedding is in association with heavier flow and greater blood loss
Menstruation (Bleeding) mechanism: • The unique features of primate females who menstruate is the existence of spiral arteries (end arteries,with no anastomosis) supplying the superficial layer of the endometrium (funtionalis layer), thus making the superficial layer of the endometrium vulnerable to ischaemia, but also facilitating hemostasis. • In the proliferative phase, the spiral arterioles grow upwards from the basal to more superficial layers of the endometrium, where a capillary network develops. • In the luteal phase, there is a marked increase in length and coiling of the spiral arterioles which will become more dilated. • Premenstrually, the endometrial glands empty secretions, the fluid from the stroma is resorbed, the endometrium shrinks (deflated), and the spiral arterioles become even more coiled up to 8 loops. At the same time, gaps appear between the endothelial cell of the spiral arterioles and the associated thin walled veins and leucocytes migrate through the gaps into the stroma which appears to be undergo disintegration.
Progesterone has a stabilizing effect and estrogen labilizing effect on lysosomes in the endometrium. The withdrawal of progesterone preceding menstruation probably causes breakdown of lysosomes and release of phsopholipase A2. This in turn causes the formation of large amounts of arachidonic acid from phospholipids in the cell wall and initiates the prostaneid cascade and the synthesis of PGF2 and PGE2 and PGI2. The sudden increase of prostaglandins, particularly PGF2 is probably responsible for the spasmodic contraction of the spiral arterioles and for menstruation. • Immediately before menstruation, the spiral arteries constrict intensely for a period of 4-24 hours and then dilate with a massive extravasation of erythrocytes into the stroma of endometrium. • Blood initially spurts from the open end of spiral arterioles but normally stops rapidly. Bleeding occurs from the coalesced blood lakes and from the torn ends of capillaries and veins, bleeding from the latter being slower and continuing longer. Approximately 75% of menstrual blood is arterial and 25% is venous. Though the proportion may change in women with menorrhagia, only about one-quarter of the total endometrium is shed; the majority involutes and is reabsorbed, as in animal species which do not menstruate. • Menstrual blood contains aggregations of erythrocytes, degraded and exhausted platelets, small amounts of fibrin and large amounts of fibrin degradation products, suggesting that the haemostatic plugs and any blood clots that may form undergo fibrinolysis and rapidly disintegrate. An excess of fibrinolytic activity in the endometrium might well impair haemostatic plug formation in the spiral arterioles and would provide a ready explanation for excessive menstrual blood loss.
Menstrual blood stoppage mechanism (Haemostasis): • Hemostatic plug function of aggregated platelets and fibrin in the spiral arterioles, which is small and incomplete compared with those in skin wounds with onion skin like allowing intermittent blood flow before complete occlusion. • Vasoconstriction of spiral arteries together with swelling of endothelial cells which completely occlude the arterioles occurs in the 2nd day of menstruation and is considered the most important mechanism controlling menstrual blood. Prostaglandins play vital part. • Re-epithelization commences from the basal glands, proceeds rapidly, and is usually completed by third or fourth day, which depends on the rate of estrogen stimulation, which in turn, depends on the rate of growth of the follicles developing in the ovaries. It starts from the region of the isthmus and cornual recesses of the ostea of the fallopian tube. Furthermore, the stromal layer contributes important autocrine and paracrine factors for growth and migration, mainly in response to injury rather than hormonal effect as hormone levels are at their nadir.
The uterus as an endocrinal organ: • The uterus is a dynamic as many endometrial products had been verified. Lipids as Prostaglandins, Thromboxanes, Leukotrienes has been identified to be secreted from stromal & endometrial cell with various functions (see below). Cytokines as Interleukin-la, lnterleukin-1B, lnterleukin-6, Interferon-g, Colony-stimulating factor-1, Tumor necrosis factor-a Leukemia,-inhibiting factor were also identified. List of other peptides secreted from endometrial cell with various functions as enzymes & enzymes inhibitors, angiogenic, vasoactive , hemostatic & growth factors includes : [ Prolactin, Relaxin, Prorenin and rennin, Endorphin, Endothelin-1,Corticotropin-releasing hormone, Fibronectin, Uteroglobin, Lipocortin-1, Parathyroid hormone-like protein, Integrins, Epidermal growth factor family [EGF / Heparin-binding EGF / TGF-a ], Insulin-like growth factor family [IGF-l / IGF-ll / IGFBPs1-6 ] , Platelet-derived growth factor, Transforming growth factor-B, Fibroblast growth factor, Vascular endothelial growth factor ].
Role of Prostaglandins: • In the proliferative phase the endometrium synthesizes equal amounts of PGF2 and PGE2 1:1, but in the luteal phase the level of PGF2 progressively increases under the influence of estradiol and progesterone 2:1 in menstrual fluid, so that vasoconstriction and platelet - aggregatory action predominates. The myometrium produces considerable amounts of PGI2 synthesized from endoperoxides produced in the endometrium which diffuse to myometrium, producing vasodilation and inhibiting platelet aggregation. • In normal menstruation, it is postulated that the PGF2 synthesized in the endometrium first produces vasoconstriction of spiral arterioles, and as a result, an increased proportion of the endoperoxides is produced from arachidonic acid by prostaglandin synthetase deviated into the myometrium, which, then produces a surge of PGI2. This surge may then diffuse back into the endometrium, producing the dilatation which follows the vasoconstriction of spiral arterioles immediately preceding the onset of menstruation.
At the end of the non-fertile cycle, endoperoxides generated from free arachidonic acid, released from membrane phospholipid stores by the action of phospholipase A2, are converted predominatly to prostaglandin F2-Endoperoxides may also be transported to the myometrium, where they are converted to prostacyclin. Prostacyclin of myometrial origin may act to prevent platelet aggregation and stimulate vasodilation within the endometrium at menstruation.
Role of Leukotrines: • They are produced predominantly by leucocytes. Excessive infiltration of the endometrium with leucocytes is seen in menorrhagia with IUDs, and the degree of menstrual blood loss is roughly proportional to the degree of infiltration. Excessive production of leukotrines is responsible for IUCD menorrhagia, if the arachidonic acid is deviated from the cyclo-oxygenase to the lipoxygenase pathway.
Enzymes & uterus: • The endometrium and cervix are sites of marked fibrinolytic activity and plasminogen activators have been demonstrated in the myometrium, endometrium and menstrual blood. The concentration of plasminogen activators in menstrual blood is maximal on the first day of bleeding and is higher in women with excess menstrual blood loss. It is also much higher in samples collected from the uterus than from the vagina, suggesting that the activators are rapidly consumed and explaining why clots may form in the vagina but rarely do so in the uterus.
Endometrial Cycle: Clinical prospective: • A normal menstrual cycle last from 21 to 35 days with 2 to 6 days of flow & an average blood loss of 20-60ml. In the extremes of reproductive age, menstrual cycles are characterized by a higher percentage of anovulatory or irregularly timed cycles. The diagnosis and management of abnormal menstrual function must be based on an understanding of the physiologic mechanisms involved in the regulation of the normal cycle. • Primary dysfunctional bleeding (PDB), including essential menorrhagia, probably results from a number of different factors, including disturbances in eicosanoid metabolism and in fibrinolytic and lysosomal enzyme systems of the endometrium. This disturbance may be primarily in the endometrium or secondary to endocrine changes originating in the ovary, pituitary and hypothalamus. • Excessive menstrual blood loss (MBL) could be due to the increased formation of lysosomes with an increased synthesis of phospholipase A2, arachidonic acid and prostaglandin at menstruation as is believed to occur in ovulatoryDUB. Marked increase in both plasminogen activators and in fibrinolytic activity plasmin in menstrual blood in case of DUB particularly with IUDs. This is reversed by anti-fibrinolytic drugs Tranexamic acid.
Interaction between thrombin formation and fibrinolysis of hemostatic plugs and the action of prostanoids including PGI2 and TXA2 and that the blood clotting fibrinolytic and prostanoid systems are closely linked. • Clinical implication of Endometrial dating: The precise nature of the histologic changes that occur in secretory endometrium relative to LH surge allows the assessment of the “normalcy” of endometrial development. Any large discrepancy (more than 2-day lag time) is termed a Luteal phase defect & has been linked to both failure of implantation & early pregnancy loss. To perform such diagnostic test, determine ovulation timing then take an endometrial biopsy 10-12 days postovulation. • Also note the clinical implication of endometrial thickness (height) by vaginal US.
The cervical cycle: • Progesterone raises the tone of the muscles of the isthmus and internal os so the cervical 'sphincter' is tighter and more competent during the luteal than during the follicular phase. • The glandular elements proliferate during the follicular phase and the epithelial cells become taller. Under the influence of oestrogens the glands actively secrete a mucus which will stretch into threads measuring more than 6.5 cm, and even 10-15 cm, at the time of ovulation. Spinnbarkeit is the basis of the thread test for estrogen in circulation. During the follicular phase the cervical mucus absorbs water and salts and, when allowed to dry, deposits crystals of sodium chloride and potassium chloride in a characteristic pattern which suggests the fronds of a fern (see diagnostic procedures).
At the time of ovulation, the secretion is so profuse that it may be noticeable as a vaginal discharge – the “ovulation cascade”. Its special character at this time makes for its easy penetration by spermatozoa. This property is related to its low content of protein. • During the luteal phase, the cervical glands become more branched and their secretion changes its physical and chemical properties. The mucus becomes more viscous and forms a more secure cervical plug. It loses, its ability to stretch without breaking and resists penetration by spermatozoa. These changes are brought about by progesterone and are related to an increase in the amount of protein in the mucus and to the presence of phospholipids.
The vaginal cycle: • The cyclical changes occur in the vaginal epithelium are better seen in smears of desquamated cells. The unstimulated vagina shows relatively small basal type cells with healthy nuclei. These and intermediate basophil forms are also seen in vaginal smears taken in the early follicular phase. The fully oestrogemc smear, evident during the late follicular phase, contains a preponderance of large cornified epithelial cells with pyknotic nuclei. These stain pink with eosin. During the luteal phase the smear shows evidence of increased desquamation, many of the cells having rolled edges, and is characterized by the reappearance of clumps of intermediate cells and the presence of leucocytes. The maturation index, which is the percentage of superficial, intermediate and parabasal cells in a vaginal smear is used as a measure of the levels of hormones in circulation. It is a useful guide but is not so precise as assaying the hormones in blood.
Cyclical changes in the tube: • The muscle of the fallopian tube behaves like the myometrium in that it shows increased movement about the time of ovulation. This is an estrogen effect. The increased cilial activity at that time. These changes are timed to propel the ovum towards the uterus. • The follicular phase is marked by slight proliferation, and this continues up to the premenstrual phase when it regresses. During menstruation there is further shrinkage and slight shedding of the surface epithelium. • The secretory activity of the tubes is also cyclical, being highest just before ovulation and in response to oestrogen. Progesterone may also play a part in this but the consensus of opinion is that hormone reduces the amount of the secretion.