Download

1 / 29
290 likes | 384 Views
Funksjonelle tilordninger i kjølvannet av Arabidopsis genomprosjektet. Per Winge stip. Biologisk inst. Organell genomer fra mitokondrie og kloroplast. Størrelse: 14 - 200 Kb. C-verdi: Mengden av nukleært DNA i en ikke replikert haploid nukleus.

E N D
Funksjonelle tilordninger i kjølvannet av Arabidopsis genomprosjektet Per Winge stip. Biologisk inst
Organell genomer fra mitokondrie og kloroplast. Størrelse: 14 - 200 Kb C-verdi: Mengden av nukleært DNA i en ikke replikert haploid nukleus. Regnes ofte i pg og er kjent for mange organismer. http://www.genomesize.com/ Definisjoner: Hva er et genom? Det nukleære genom. Størrelse 30-100,000 Mb Alt nukleært DNA i celle kjernen.
mRNA blir revers transkribert til cDNA og deretter sekvensert. Proteom Det komplette sett av proteiner kodet i fra et genom Proteomics Studier av proteiner uttrykk fra et genom. Transkriptom Det komplette sett av RNA som blir transkribert i fra et genom. Eukromatin De gen rike områdene i fra et genom. Heterokromatin Kompakte, gen fattige områder av et genom. BAC ”Bacterial Artificial Chromosome”. Et ”kunstig” bakterielt kromosom som benyttes som kloningsvektor. EST ”Expressed sequence tag”. En kort DNA sekvens som kommer i fra et mRNA.
Prokaryot Enkeltcellet organisme uten cellekjerne og med enkel intern struktur. Eukaryot En organisme (enkeltcellet / multicellulær) som har en kompleks intern struktur inkludert en celle kjerne. Protist Eukaryoter som ikke kan grupperes under planter, dyr eller sopp/gjær. Ortholog Gen som forblir ”uforandret” også etter en arts dannelse. Har vanligvis samme lokus i beslektede organismer. Paralog Et gen som har oppstått som følge av en gen duplikajon.
Hvordan er slektskapet mellom levende organismer? Sammenligninger av ribosomal RNA og konserverte proteiner har vist at prokaryote organismer kan deles in i to hovedgrupper: bakterier og archaea bakterier. Levende organismer kan derfor deles inn i tre domener: Bakterier, Archaea og Eukaryoter. Archaea bakteriene kan videre sub-inndeles i Crenarchaeota og Euryarchaeota. Flere av Archaea bakteriene er såkalte ekstremofile. Dvs. de lever under ekstreme betingelser, høy temperatur eller i omgivelser med høyt saltinnhold, lav Ph osv.
De tidligste eukaryote cellene oppsto sansynligvis som en hel celle fusjon / symbiose mellom en archaea bakterie og en eubakterie, (1.6 – 2.1 milliarder år siden). Dette ga opphavet til en amitokondriell (uten mitokondrie) eukaryot celle. De mest primitive eukaryote organismene, archeaprotister som Giardia (Diplomonader) og trichomonader (Parabasalidea), mangler mitokondrier, nukleoli, peroxisomer, og har et relativt primitivt cytoskjellet. De beveger seg ved hjelp av flageller og flimmerhår.
Slektskapet mellom ulike grupper av protister er fremdeles uklart men molekylær fylogenetiske studier begynner å gi noen svar. Amitokondrielle protister er de mest avvikende og peker seg ut som den gruppen som oppsto først. Hypotese: Opphavet til mitokondrielle eukaryoter. Amitokondrielle protister begynte å leve i symbiose med en alfa-proteo bakterie, endosymbiont teorien. Gener fra bakteriellt genom ble enten deletert eller overført til den eukaryote cellekjernen. En viktig konsekvens av denne symbiosen er at de tidlige eukaryote cellene, som mest sannsynlig var begrenset til en anaerob metabolisme, nå kunne foreta aerobisk respirasjon. Senere har enkelte protister som lever som animalske parasitter mistet mitokondriene og blitt sekundært amitokondrielle, for eksempel ulike mikrosporidia.
Sammenligning av komplette genom i fra eubakterier, archea bakterier og eukaryoter viser at eukaryote proteiner som inngår i prosesser som replikasjon av DNA, transkripsjon av mRNA, translasjon samt organisering av endoplasmatisk retikulum er nærmere beslektet med proteiner fra archaea enn fra bakterier. Mitokondriet oppsto sansynligvis som et resultat av en symbiose mellom en alfa-proteobakterie og en tidlig eukaryot celle Eukaryote proteiner som inngår i metabolisme, detoksifisering og regulering av ionebalanse er nærmere beslektet med bakterielle proteiner.
Viridiplantae inkluderer ulike grønnalger samt høyere planter (landplanter). Viridiplantae ser ut til å ha oppstått som en endo-symbiose mellom en cyanobakterie og en primitiv eukaryot med mitokondrier. Rester av denne symbiosen finner en i dag i kloroplastene og slektskapet mellom kloroplast proteiner og cyanobakterie proteiner er nært. En rekke av de opprinnelige kloroplast genene (cyanobakterie genene) er senere overført til kjernen. I Arabidopsis thaliana er over 1000 proteiner (av ~ 25,000 totalt) nært beslektet med proteiner en finner i cyanobakterier. Prasinophytes Chlorophyceae Trebouxiophyceae Ulvophyceae Chlorokybales Klebsormidiales Streptophyta Zygnematales Charales Coleochaetales Embryophytes (landplanter)
De mest primitive landplantene (embryofyta) er sannsynligvis levermosene (Marchantiofyta) og det er antatt at de har eksistert i over 480 millioner år. Frø planter
Vaskulære planter oppsto for ca. 420 millioner år siden. Inndeling av frøplanter på grunnlag av molekylære data. De første frøplanter oppsto for rundt 380 millioner år siden og blomster planter har kun eksistert i ca 200 millioner år. En frøbladet og to frøbladet planter oppsto for ca 120-150 millioner år siden. Eudicots kan videre sub-inndeles i to store familier, ”Asterids” (f. eks. tomat) og ”Rosids” (f. eks. Arabidopsis). Disse to gruppene oppsto for ca 90 millioner år siden.
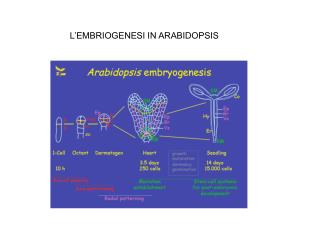
Arabidopsis thaliana, vårskrinneblom Tilhører Brassicaceae familien og ligger under ordenen Capparales. Nært beslektede planter er f. eks. raps og kål. Arabidopsis thaliana har i flere år vært en av modell organismene innen plante biologi. Dette er på grunn av flere faktorer. Arabidopsis thaliana har en kort livssyklus, ca. 6 uker i fra spiring til den utvikler modne frø. Den er enkel å dyrke, er liten av vekst (ca. 30 cm høy), og trenger lite dyrkningsplass. Arabidopsis thaliana er en selv-pollinator, dvs. en trenger ikke manuelt å pollinere blomstene (ev. være avhengig av insekt pollinatorer). Kryss-pollinering er også relativt sjelden. Det er derfor lett å opprettholde ”rene linjer”. Den lar seg lett transformere med jord bakterien Agrobacterium tumefaciens som kan brukes til å overføre ulike gen konstruksjoner.
Arabidopsis thaliana, vårskrinneblom Arabidopsis er ”diploid” og har et genom på ca. 125 mega baser. Inneholder lite repetert DNA og i over 10 år har det eksistert relativt gode genetiske kart. Det finnes et stort utvalg av Arabidopsis mutanter som kan fås i fra frøbanker / ”Stock center” i USA og England. T-DNA “knockout” linjer. Hvor T-DNA fra Agrobakterium vektor er satt inn i kjent og ukjente gener og inaktivert disse. (Stort potensiale for den som vil drive med “reverse-genetics”. Over 10,000 full lengde cDNA kloner er tilgjengelige. BAC kloner fra hele genomet er tilgjengelige.
Arabidopsis ble i Desember 2000 den første planten hvor ”hele” genomet (minus enkelte centromer regioner) ble oppklart / sekvensert. Arabidopsis genomet består av 5 kromosom som varierer i fra 17.5 til 29.1 mega baser. Sekvenseringen av Arabidopsis ble foretatt av et internasjonalt konsortium ”The Arabidopsis Genome Initiative” i tidsrommet 1996-2000. Mesteparten ble sekvensert de siste 2 årene.
På grunn av at gode genetiske kart eksisterte for Arabidopsis og at genomet var relativt lite ble en ”mapping” basert sekvenseringsstrategi benyttet. Dvs. det ble laget et genomisk BAC bibliotek, hvor overlappende kloner ble identifisert ved hjelp av RFLP analyser og hybridisering eller ved PCR av ”sequence-tagged sites” (STS) og Southern blotting. BAC kloner ble shotgun sekvensert. BAC klonene (ca. 50-100 kb.) ble enkeltvis kuttet opp i småbiter og klonet over i plasmid vektorer (1-3 Kb) og deretter sekvensert. Overlappende sekvenser ble deretter satt sammen til sekvens av hele BAC klon var komplett.
Resultatet fra sekvenseringen viste at Arabidopsis thaliana genomet var rundt 125 mega baser og inneholdt rundt 25,000 gener. Dvs. flere gener enn man fant i de to invertebrate organismene; nematoden Caenorhabditis elegans (ca. 19,000 gener) og i bananflua Drosophila melanogaster (ca. 13,500 gener). Selv om sekvensen til genomet er kjent er det fremdeles mye arbeid som gjenstår. Å sette sammen alle exon i et gen korrekt, samt å finne start / stopp og beskrivelse av protein er ikke trivielt. Denne prosessen, som kalles for annotering, er i mange tilfeller gjort av dataprogram og er ofte unøyaktig. Etter at dataprogrammet har funnet et mulig gen og korresponderende protein blir det sjekket mot gen / protein databasene: GenBank NCBI), EMBL. Protein domene struktur kan analyseres gjennom Pfam databasen (Sanger Centre). Videre annotering av gen / protein. GeneOnthology: Grupperer protein i henhold til funksjon, Metabolisme, Celle syklus regulering, DNA reparajon osv.
Eksempel på analyse av et protein Protein sekvens. >gi|2352084|gb|AAB68776.1 MDHNSPKSRRSRKPEPKPDIYSTFVVHSDSDSDQGRDRDKRKAKPEEDENVDLYATMVYKGDSDGEGEED DDDDSMLPPLLKRLPKDFGGGASLDYDDDDGDESGDFGTMIVKTDRSSHSKKNSPYSSKPRMGVSPRRRA RGGDEESSDEEDEEEDDDDDDGDYGTFVVKSKDKKGKKKDKEIDMTTMGRAVASMQKSNFGGKTRKLDPS SSSSKLHGEDNRKMQQQNSKMSTTSLPDSITREDPTTKYEFLNELGKGSYGSVYKARDLKTSEIVAVKVI SLTEGEEGYEEIRGEIEMLQQCNHPNVVRYLGSYQGEDYLWIVMEYCGGGSVADLMNVTEEALEEYQIAY ICREALKGLAYLHSIYKVHRDIKGGNILLTEQGEVKLGDFGVAAQLTRTMSKRNTFIGTPHWMAPEVIQE NRYDGKVDVWALGVSAIEMAEGLPPRSSVHPMRVLFMISIEPAPMLEDKEKWSLVFHDFVAKCLTKEPRL RPTAAEMLKHKFVERCKTGASAMSPKIEKSRQIRATMALQAQSVVAPSLEDTSTLGPKSSEELGITVPSK PPQNSTEAPLTSTLNRQHITGNTVLAGEGGDFGTMIVHGEDETEESDSRSQLVREKESSSSQFEGVPREF PGEELPDSWIHDKKKPPAIDLPVEASISQSMQASSSHEHRTKLHNIAGTQMEGGSDASGSTLKNETVGRK AFALQDKLWSIYAAGNTVPIPFLRATDISPIALLSENMIGGMQQDGNGTVAVEALQELFTSSDPQSKKGR RGQNEMPLPPSVYQRLTTSSSLMNLAQVLAYHRACYEEMPLQELQATQEQQTIQNLCDTLRTILRL BlastP analyse. Score E Value Sequences producing significant alignments: gi|2352084|gb|AAB68776.1| (U96613) serine/threonine kinase ... 1324 0.0 gi|12061243|gb|AAG45491.1| (AY013245) 36I5.3 [Oryza sativa] 659 0.0 gi|3114674|gb|AAC15972.1| kinase [Dictyostelium discoideum] ... 282 1e-74 gi|7302511|gb|AAF57595.1| CG7097 [Drosophila melanogaster] ... 272 2e-71 gi|12643529|sp|O00506|ST25_HUMAN SERINE/THREONINE PROTEIN K... 271 2e-71
Kun 10-15 % av genene i Arabidopsis thaliana er annotert og karakterisert eksperimentelt (ikke maskinelt). Over 30% av genene er uklassifiserte, hypotetiske, ”ukjente”.
Sammenligning av Arabidopsis genom med ulike andre kjente genom
Sammenligningen mellom Arabidopsis thaliana genomet og de kjente genomene fra cyanobakterier, gjær, bananflue og nematode, viser bl. a. Antall gener som er involvert i cellulær kommunikasjon og signaloverføringer en hel del høyere i multicellulære organismer. Planter har utviklet en rekke unike transkripsjonsfaktorer som ikke finnes i animalia / fungi. En rekke av Arabidopsis proteinene som er inkludert i kategoriene energi og metabolisme har høy homologi med bakterielle proteiner (cyanobakterium). Arabidopsis inneholder rundt 11000 typer genfamiler eller ”singletons”, noe som er relativt likt det vi finner i Drosophila og C. elegans. Arabidopsis og planter generelt har utviklet unike signaltransduksjonsspor som en ikke finner i animalia og gjær. Arabidopsis har f. eks. ikke komponenter som inngår i de kjente signalsporene: - Reseptor tyrosin kinaser / ras ”pathway” - Nukleære steroid hormon reseptorer - Wingless / wnt - Notch / lin12 - JAK / STAT signal sporet
Arabidopsis genomet inneholder en rekke store duplikasjoner og gener som er duplisert i tandem. Dette betyr at mange gener har en eller flere paraloger. Omfanget av disse duplikasjonene er så store at enkelte har foreslått at forløperen til Arabidopsis thaliana var tetraploid og at den senere har blitt ”diploidisert”.
Chr II T17A5 16 Chr III Gene duplications Nærmere studier av disse duplikasjonene tyder på at de skyldes fem (eller flere) store duplikasjoner (ev. genom duplikasjoner) som skjedde for ca. 50, 100, 140, 170 og 200 millioner år siden. Disse duplikasjonene korresponderer ganske bra med tidsepokene når f. eks “monocots” (en frøbladet) og “dicots” (to frøbladet) oppsto (ca. 120 - 150 millioner år siden) eller når splittelsen mellom ”rosids” og ”asterids” oppsto (ca. 90 millioner år siden). Med bakgrunn i denne informasjonen kan en nå ha en viss oversikt over hva en kan forvente å finne i andre planter. Informasjonen i fra Arabidopsis genomet viser også hvor dynamisk plante genomene er. Store rearrangeringer av genomene (duplikasjoner og delesjoner) er vanlige og ekspansjon av genfamilier gjenom tandem duplikasjoner er vanlig.
F4I1 19 F4I1 18 ARAC9 II Thaumatin homologues Ascorbate peroxydase T19K4 140 T19K4 130 T19K4 110 T19K4 100 T19K4 70 T19K4 60 T19K4 50 T19K4 40 T19K4 30 T19K4 20 T19K4 10 ARAC6 IV Histidine kinase ARAC1 VPS35 II T17A5 16 PAD2- like MS5- like AtCrn ARAC11 VPS35 FH3 III Gene duplications Putative anti- fungal proteins Anther-specific proline rich proteins VPS35 CLV1 ARAC5 I TUB5 ARAC4 I AtCrn ARAC2 0 10000 20000 30000 40000 50000 V Chromosome Eksempel på gen duplikasjoner i Arabidopsis thaliana
Andre observasjoner: Gener som inngår i DNA replikasjon og DNA reparasjon er forholdsvis nært lokalisert på kromosom. Tilfeldighet eller har plantegenomet fremdeles en operon lignende struktur / gener samlet i større regulon. Det er indikasjoner på at gener er lokalisert i ”funksjonelle” gen “cluster”, DNA reparasjon, planteforsvar, metabolske spor osv. Å plassere gener som inngår i samme prosesser fysisk nær hverandre kan av og til være fordelaktig. Ko-regulering av gen transkripsjon for eksempel. Gener en ikke finner i Arabidopsis men som eksisterer i gjær og animalia: - Cdc42, regulator av cellepolaritet og signaltransduksjon - Cdc25 fosfotyrosin fosfatase, celledelings-regulator - Bcl2, apoptose regulator - DH domene proteiner, Rho guanin utbyttingsfaktorer - Ras GTPase (signaltransduksjon) - Rel / NF-kappaB family - Septin GTPase (celle deling)
Hva nå? ”Big ideas from a small plant” I de neste 1-3 årene: Man vil prøve å generere knockout mutanter av de fleste genene i Arabidopsis. Disse metodene er basert på at T-DNA ev. et transposon setter seg inn i et gen og slår ut funksjonen. Over halvparten av genene er tilgjengelige som T-DNA knockout allerede i dag. riboflavin kinase T-DNA knockout
Gener som ikke lar seg slå ut ved standard T-DNA eller transposon knockout vil bli forsøkt nedregulert ved hjelp av RNA interferens (RNAi). RNA avhengig RNA polymerase
Stor skala analyser av genekspresjon ved hjelp av DNA mikroarrays er allerede i gang og vil være et nyttig verktøy for å få oversikt over hvilke gener som er uttrykt i de enkelte vev, indusert og ikke indusert.
Kryssing av to gjær stammer som uttrykker to proteiner som er fusjonert til “bait” og “pray” proteiner. Vellykket protein- interaksjon setter i gang transkripsjon av seleksjonsgener som gjør at gjær kan dyrkes på et seleksjonsmedium. Start mRNA syntese Seleksjons gen Hvor går Arabidopsis forskningen de neste 3-10 årene. Stor skala analyse av protein interaksjoner, ved bruk av gjær to hybrid system. Analyse av protein interaksjonsnettverk
Bestemme tre dimensjonale strukturer av alle grupper av proteiner. Med økende antall kjente protein strukturer er det mulig at de fleste 3D strukturer kan modelleres med stor grad av nøyaktighet allerede om 3-5 år. Hvor går Arabidopsis forskningen de neste 3-10 årene. Lage komplette samlinger av full lengde cDNA. Pr. dato er over 10,000 cDNA kloner tilgjengelige. Uvurderlige for protein interaksjonsstudier. Bestemme biologisk funksjon for ”alle” proteiner. Karakterisere cis regulatoriske sekvenser i alle gener. Dvs. stor skala promoter analyser. Utvikle bioinformatikk verktøy, visualisering og modelleringsmetoder som integrerer ”all” biologisk informasjon fra Arabidopsis. ”The virtual plant”.