Download
1 / 38
380 likes | 521 Views
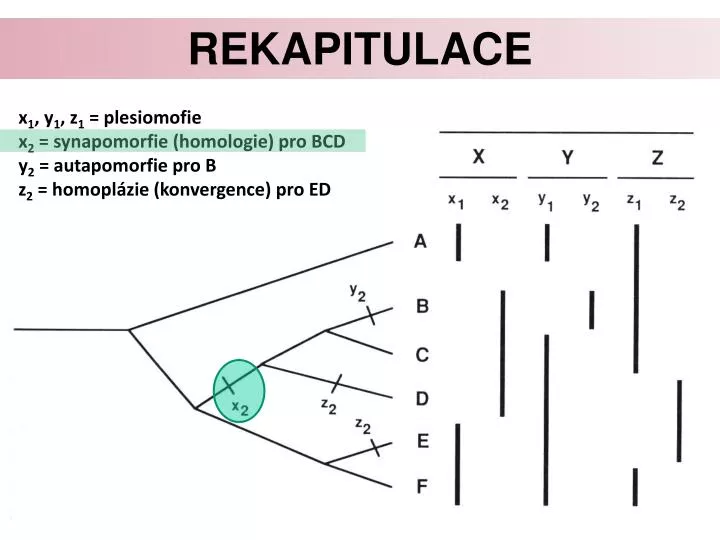
Rekapitulace. x 1 , y 1 , z 1 = plesiomofie x 2 = synapomorfie (homologie) pro BCD y 2 = autapomorfie pro B z 2 = homoplázie (konvergence) pro ED. rEKAPITULACE. Jak získáváme znaky pomocí sekvenace unikátních lokusů. Sekvenace lokusu u taxonů, které chceme studovat 1. Přednáška.

E N D
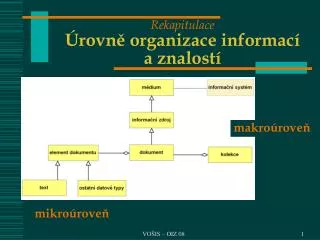
Rekapitulace x1, y1, z1 = plesiomofie x2 = synapomorfie (homologie) pro BCD y2 = autapomorfie pro B z2 = homoplázie (konvergence) pro ED
rEKAPITULACE Jak získáváme znaky pomocí sekvenace unikátních lokusů Sekvenace lokusu u taxonů, které chceme studovat 1. Přednáška Stažení homologních sekvencí od relevantních taxonů z databáze 2. přednáška Tvorba alignmentu 3. přednáška Bude náplní prvního praktika tento pátek 9:50, B5
alignment Mutliple sequence alignment (MSA) Kontrolní otázka: Co v alignmentu představuje jeden znak?
Ostatní metody získávání molekulárních dat Pokud bychom měli k dispozici kompletní genomové sekvence organismů, které chceme studovat, žádné takové metody bychom nepotřebovali. Jenže je nemáme a jejich sekvenování je stále neprakticky drahé. Následující metody umožňují rychle a často levně získat dostatečné množství dat pro otázky, které v molekulární taxonomii řešíme.
DNA-DNA hybridizace Kvantifikace reasociace • Stanovit střední teplotu tání homoduplexů • Tm= (TmA + TmB)/2 • Stanovit teplotu tání heteroduplexu • Vypočítat pokles ΔTm • ∆Tm=((TmA + TmB)/2) - Tms • Vypočítat p (podíl rozdílných nukleotidů) • p = ΔTm . 0,01 (0,015) Kontrolní otázka: Co u téhle metody představuje jeden znak?
DNA-DNA hybridizace Výhody • Multilokusová (totilokusová) metoda • Použitelná na nejvzdálenější taxony Nevýhody • Distanční metoda • Experimentálně dosti náročná • Vyžaduje větší množství DNA (mg) • Náchylná k artefaktům (repetitivní DNA) • Experimentální náročnost N x N
IZOENZYMOVá analýza • Isozymy– enzymy vykazující stejnou nebo podobnou aktivitu. • Alozymy– podmnožina isozymů, alelické formy téhož enzymu (v jednom lokusu) • Zymodem -taxonomická jednotka vymezená na podkladě isozymového vzoru • Zymogram - rozložení zón a tedy i jednotlivých enzymů na elektroforetogramu
IZOENZYMOVá analýza Princip Detekce elektroforetickou mobilitou se lišících forem enzymů kódovaných různými alelami určitého genu. Jednotlivé enzymy se detekují prostřednictvím jejich specifické aktivity, většinou pomocí spřažené enzymatické reakce s barevným produktem.
IZOENZYMOVá analýza Biologická podstata • Velké množství genů pro enzymy je polymorfních (nejhojnější alela <90 %) • Velká část tohoto polymorfismu představuje polymorfismus vnitropopulační (obvykle 15 % genů je polymorfních, od 0-50 %) (málo ptáci!) • Alozymy se liší sekvencí aminokyselin, značná část změn se projeví změnami elektromobility.
IZOENZYMOVá analýza Detekce Princip: Tetrazoliové soli jsou redukovány na modrý formazan Chromogenní substrát: methyl thiazolová modř(MTT) přenašeč elektronů: phenasin methosulfát (PMS) Glukozofosfát isomeráza GPI GPD glukonolakton-6-fosfát fruktozo-6-fosfát glukozo-6-fosfát PMS NADP NADPH NADP MTT formazan fruktozo-6-fosfát, NADP, MgCl2 ,MTT, PMS, glukozo-6-fosfát dehydrogenáza
IZOENZYMOVá analýza Výhody • Kodominance • Množství polymorfismu je vhodné pro vnitropopulačnístudie • Znaková metoda • Nízká cena analýzy a cena vybavení Nevýhody • Nemusí být selekčně neutrální (mikrosatelityjsoulepší) • Konvergence (i různé alozymy mohou mít shodnou elektromobilitu) • Maximální divergence 60-70 % • u žab na úrovni rodu či spíše druhů • u savců na úrovni podčeledí
IZOENZYMOVá analýza Využití • Studium populační struktury • rozmnožovací struktura (např. panmixie) • migrace • genetická příbuznost populací • Studium druhové diversity • kryptické (sibling) druhy (sym/alopatrické) • determinace známých druhů • Fylogeneze
Mikrosatelity Mikrosatelity- (nazývané i STR - Short Tandem Repeat) jsou krátké sekvenční motivy (dinukleotidy až hexanukleotidy) vyskytující se na některých místech genomu v mnoha tandemově uspořádaných kopiích o celkové délce až 150 nukleotidů. Zpravidla existuje velmi značný vnitropopulační polymorfismus v počtech kopií daného mikrosatelitu a tento polymorfismus a mutabilitu (okolo 0,1 % mutací za generaci) můžeme snadno studovat například PCR amplifikací daného lokusu.
Mikrosatelity Postup při analýze • najít vhodný lokus (genomová knihovna a hybridizace) • připravit PCR primery ohraničující daný lokus • amplifikovat lokus • elekroforéza amplifikovaných fragmentů
Mikrosatelity Využití metody • vnitrodruhové studie či příbuzné druhy • kodominance znaků – populační genetika • modernější a přesnější varianta alozymové analýzy • možno analyzovat velké množství vzorků • nákladné pouze zavedení metody pro nové organismy
Restrikční analýzy • Izolace DNA • Nastříhání DNA pomocí restrikčních endonukleáz • Velké množství endonukleáz, možno použít i jejich kombinace • Problémy s přílišnou komplexitou vzorů • Snížení komplexity výchozí DNA • Kvalita separace fragmentů • Specifičnost detekce zón EcoRI
Restrikční analýzy RFLP bakteriální DNA Komplexita bakteriálního genomu je nízká, a proto vzniká i méně komplexní vzor, ze kterého je možno odečíst jednotlivé pruhy Celková DNA po naštěpení rozdělena na PAGE a obarvena EtBr
Restrikční analýzy RFLP organelové DNA Izolovaná cpDNA naštěpena enzymy, rozdělena na agarose a obarvena EtBr.
Restrikční analýzy mtRFLP – krok 1 Celková DNA rozštěpena, rozdělena na agarose, obarvena EtBr
Restrikční analýzy mtRFLP – krok 2 – Southern blot
Restrikční analýzy mtRFLP – krok 2 DNA z agarosy přeblotována na nylon, hybridizovánas mtDNA sondou a detekována autoradiograficky.
Restrikční analýzy VNTR – Variable Number of Tantem Repeats Využívá polymorfismus v počtu kopií tandemových repetic – minisatelitů (10-60 nt). Tento polymorfismus je velmi variabilní i mezi jedinci téhož druhu. Celkovou DNA naštípeme restrikční endonukleázou štěpící mimo minisatelit. Naštípanou DNA přeblotujeme na membránu a hybridizujeme se značenou próbou buďto proti minisatelitu (pokud chceme zviditelnit všechny lokusy) nebo nebo proti minisatelitua unikátní sekvenci v sousedství (pokud chceme zviditelnit jeden lokus, na obrázku).
Restrikční analýzy Výhody • Relativní jednoduchost a láce • Velké množství dat • Slušná reprodukovatelnost • Kodominance znaků • Multilokusový charakter • Selekční neutralita znaků
Restrikční analýzy Nevýhody • Potřeba většího množství DNA • Potřeba kvalitnější a čistší DNA • Spíše pro příbuzné druhy • Někdy špatně čitelné elektroforetogramy – nutnost snižovat komplexitu DNA • Znaky nemusí být nezávislé • Spíše distanční metoda
SINE Short INterspersed repetitive Elements Retroposony derivované z tRNA, případně 7 SL RNA (Alu). Velikost: 70-500 PB. Vytvářejí značnou část eukaryotického genomu, často až 104 kopií na genom. Výhoda pro molekulární fylogenetiku existuje jen malá pravděpodobnost, že by se u dvou nepříbuzných organismů vložily do stejného místa a je téměř vyloučeno, že by se bezestopy z daného místa vystřihly.
SINE Postup • Najít nový SINE element (in vitrotranskribce celkové DNA) • Naklonovat úseky genomu obsahující daný SINE (genomová knihovna) • Vytipovat SINE lokusy které jsou polymorfní u studovaného taxonu (hybridizace nebo PCR) • PCR amplifikace vybraného lokusu • Ověření absenci SINE sekvenací získaných krátkých PCR fragmentů
SINE PCR produkty EtBr Hybridizace se SINE probou Hybridizace s okolní unikátní sekvencí
SINE Využití • použitelná pro vyšší taxony (maximálně však 75-100 milionů let • málo znaků – není možno počítat délku větví • unikátní události malá pravděpodobnost homoplázií • vždy nutno ověřit absenci SINE v daném lokusu minimálně amplifikací, raději sekvenací
RAPD RandomAmplifiedPolymorphicDNA • Amplifikace pomocí „náhodných“ primerů • Různé metody zvyšování počtu znaků • Kombinace primerů, restrikce enzymy, semiselektivníprimery (transposomy)
RAPD Výhody • Cena • Rychlost • Univerzálnost • Malé množství materiálu • Nižší požadavky na kvalitu DNA • Obrovské (libovolné) množství dat • Kvalita elektroforetogramů
RAPD Nevýhody • Neprodukuje věrohodné znaky (mnohofalešných homologií), při analýze je třeba převést na genetické vzdálenosti • Použitelné jen u relativně příbuzných druhů • Chybí kodominance • Interference znaků • Vliv kontaminující DNA • Nízká reprodukovatelnost PCR
AFLP Amplified Fragment Length Polymorphism
AFLP kapilárová elektroforéza standard molekulární váhy amplifikované fragmenty
AFLP Výhody • Vysoká reprodukovatelnost výsledků • Velké množství znaků • Kodominance znaků • Minimum falešných homologií • Lze použít znakově orientované metody Nevýhody • Nákladnost kitů i přístrojového vybavení • Složitost metody • Spíše pro příbuzné druhy