Download
1 / 24
240 likes | 384 Views
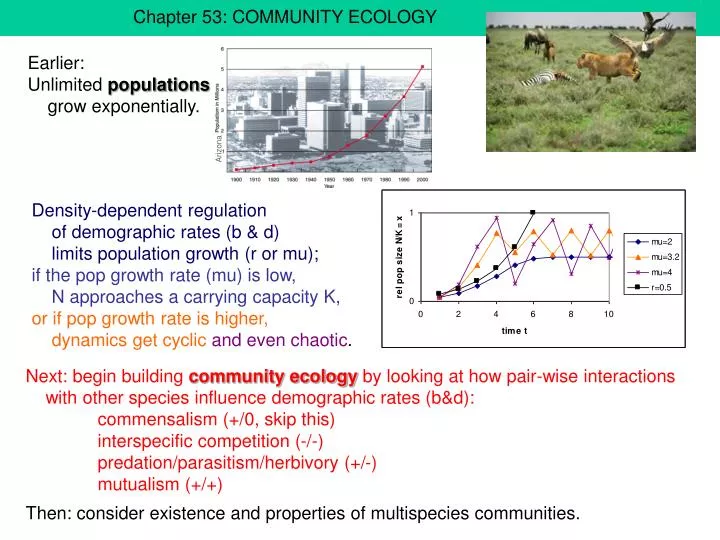
Arizona. Chapter 53: COMMUNITY ECOLOGY. Earlier: Unlimited populations grow exponentially. Density-dependent regulation of demographic rates (b & d) limits population growth (r or mu); if the pop growth rate (mu) is low, N approaches a carrying capacity K,

E N D
Arizona Chapter 53: COMMUNITY ECOLOGY Earlier: Unlimited populations grow exponentially. Density-dependent regulation of demographic rates (b & d) limits population growth (r or mu); if the pop growth rate (mu) is low, N approaches a carrying capacity K, or if pop growth rate is higher, dynamics get cyclicand even chaotic. Next: begin building community ecology by looking at how pair-wise interactions with other species influence demographic rates (b&d): commensalism (+/0, skip this) interspecific competition (-/-) predation/parasitism/herbivory (+/-) mutualism (+/+) Then: consider existence and properties of multispecies communities.
The individualistic hypothesis of community structure was enunciated by H. A. Gleason in the early 1900s. It depicted the plant community as a chance assemblage of species in the same area simply because they happen to have similar abiotic requirements - for example, for temperature, rainfall, and soil type. The interactive hypothesis was advocated by F. E. Clements in the early 1900s. … the community as a group of closely linked species, locked into association by mandatory biotic interactions that cause the community to function as an integrated unit - as a superorganism. In most actual cases where there are gradients of environmental variation, the composition of plant communities seems to change continuously, with species more or less independently distributed. … support the view of plant communities as loose associations without discrete boundaries. A community is any assemblage of populations in an area or habitat. How can we account for the species found together as members of a community? Two different views on this question emerged among ecologists.
incorporate Intraspecific competition w/ density-dependent -FB on growth rate r The logistic growth model: dN/dt = N [rmax (1 - N/K)] The Lotka-Volterra interspecific competition model: dN1/dt = N1[r1max (1 - N1/K1- a12 N2/K1)] dN2/dt = N2[r2max (1 - N2/K2- a21 N1/K2)] The exponential growth model: dN/dt = N r How can we add a little more realism to this model by incorporating Interspecific Competition? Suppose we can measure the impact of 1 individual of species 2 on the pop growth rate of species 1 relative to the impact of 1 additional individual of species 1? Call this rate of substitution the ‘interspecific competition coefficient’ a12 (the intraspecific coefficients a11 & a22 are implicitly = 1) If we ‘do the math’ we find stable coexistence of both species 1 & 2 only if a12< 1 & a21< 1,meaning: intraspecific competition > interspecific competition, implying: diff species use resources differently {different niches}. Otherwise, expect competitive exclusion of one of the species.
Competitive exclusion is often found in simple lab experiments where 2 diff species are forced to scramble for one limiting resource. (it’s difficult to est stable pairs in simple environments w/ exploitation competition). A classic illustration is from Gause’s exp’s w/ paramecia competing for bacteria in chemostats (w/ fixed input of food). When grown separately each species does fine (note: P. aurelia has smaller r but larger K) When grown in competition, P. aureliadrivesP. caudatum out of the system Competitive exclusion is expected from details of L-V competition model, but there is a simpler interpretation (from D.Tilman): Note that P. aurelia has a bigger carrying capacity K than P. caudatum. This means that each P aurelia can survive & reproduce w/ less resources: ((1/KPa)th of the resource base) < ((1/KPc)th of the resource base). The growing pop of P.aurelia can drive resources below the minimun required for individual P. caudatum to reproduce.
Chthamalus tolerates a broad range of exposure to air (broad fun. niche), but is a poor sumo wrestler. Balanus cannot tolerate long exposure to air (narrow fun. niche), but in lower inter-tidal, is very aggressive - pries off Chthamalus. A classic illustration of competitive exclusion in the field (involving interference/aggression) from Connell, Fig 53.2 Even though Chthamalus is concentrated primarily on the upper strata of rocks, when ecologist Joseph Connell removed Balanus from the lower strata, the Chthamalus population spread into that area. Thus, Chthamalus could survive lower on the rocks than where it is generally found, were it not for competition from Balanus . Its realized niche is only a fraction of its fundamental niche. Note: coexistence results from tradeoffs; each species better at something.
Prospects for 'competitive exclusion' treatment to control salmonellas and other foodborne pathogens in poultry.Mead GC. Veterinary Journal 159: (2) 111-123 MAR 2000. In newly hatched chicks, the rapid establishment of an adult-type intestinal microflora, via the oral route, produces almost immediate resistance to colonization by any food poisoning salmonellas that gain access to die rearing environment.Exploitation of the 'competitive exclusion' effect is now an accepted part of the overall strategy by which poultry-associated salmonellas are being controlled in some countries. My enemy’s enemy is my friend! - recall vaginal yeast infection story.
The Galápagos finches provide a good example of character displacement in beak sizes and, presumably, in the seeds that they can eat most efficiently. Allopatric populations {in different places} of Geospiza fuliginosa and G. fortis have beaks of similar size, but on an island where both species occur, a significant difference in beak depth has evolved. It is a challenge to develop an appropriate null model for differences expected w/o past competition & character displacement. There are two possible outcomes of competition between species w/ identical niches: either the less competitive species will be driven to local extinction, or one of the species may evolve to use a different set of resources. This differentiation of niches is called resource partitioning. We can think of resource partitioning as "the ghost of competition past“ - circumstantial evidence of earlier interspecific competition resolved by the evolution of niche differentiation. Niche differentiation is often evident from morphological character displacement.
It is apparent that predation influences the distribution and abundance of prey species - most obviously by killing some and scaring the rest. Predators can actually increase biodiversity, by disturbing competitive exclusion: A keystone predator blockscompetitive exclusion by preferentially killing the most abundant (dominant) competitor. (R. Payne, Fig 53.14) w/o starfish eating mussels, mussels displace everything else
Parasites & disease influence the distribution and abundance of organisms http://www.hhmi.ucla.edu/C168/week6/lecture1/lecture1.html The African trypanosome, Trypanosoma brucei, is an important pathogen of humans and livestock. … the parasite and the subsequent disease in cattle renders much of Central Africa refractory to raising high-yielding beef and dairy cows; indirectly, this has the most profound effect on the human economy and nutrition in the area. {& explains why cows haven’t yet displaced wildlife} The reservoir hosts are wild game animals (antelope and deer). Humans become infected when they invade the territory of the tsetse fly. Consider chestnut blight & Dutch elm disease. … the parasite can change its surface coat at about the same frequency as the host can mount an immune response to new antigens.
In ecology, as w/ gravity, ‘everything is connected to everything else’ - the trick is figuring out which interactions are ‘trivial’ & can be ignored. Chain reactions linking acorns to gypsy moth outbreaks and Lyme disease risk.Jones et al. 1998. Science 279:1023-1026. Borrelia burgdorferi Humans (Lyme disease) - + + http://www.ecostudies.org/research/reports/ostrept1.html + + deer ticks mice + deer - + acorns gypsy moths + - oak trees
Competition between the gypsy moth, Lymantria dispar, and the northern tiger swallowtail, Papilio canadensis: interactions mediated by host plant chemistry, pathogens, and parasitoids.Redman AM, Scriber JM - OECOLOGIA 125: (2) 218-228 OCT 2000
Redman & Scriber - OECOLOGIA 125: (2) 218-228 OCT 2000 … interspecific competition among insect herbivores … probably involves indirect, or "apparent", competition more frequently than direct battles over food … Typically, indirect competition between species occurs when the activity of one species induces a chemical response in its host plant, affecting other species feeding on the same plant. Examples of indirect competition mediated by pathogens have been reported for a great many species … but not, as far as we know, for herbivorous insects. … predators and parasitoids may also mediate indirect competition … The attraction of polyphagous natural enemies to visual cues or … host damage can negatively affect neighboring herbivores.
Redman & Scriber - OECOLOGIA 125: (2) 218-228 OCT 2000 We investigated interspecific competition in a study system involving the northern tiger swallowtail and the gypsy moth, feeding on quaking aspen. The northern tiger swallowtail, Papilio canadensis is native to North America. … P. canadensis … typically exhibits a strong preference for quaking aspen, Populus tremuloides … Females … oviposit on newly expanded leaves during the first few weeks of June, and larvae grow throughout the summer, pupating in mid- to late August. Quaking aspen, which supports a large number of herbivores, is defended primarily by the phenolic glycosides salicortin and tremulacin, and secondarily by tannins. when leaf tissue is removed … refoliation replaces nearly 90% of damaged foliage with leaves that tend to be rich in phenolic glycosides. Damaged leaves themselves also tend to contain elevated levels of these compounds.
Redman & Scriber - OECOLOGIA 125: (2) 218-228 OCT 2000 The European gypsy moth, Lymantria dispar was brought to Massachusetts from France in 1868, and has since radiated … over most of the eastern United States. Gypsy moth outbreaks are ecologically dramatic events, and it is difficult to imagine that many species remain unaffected … Gypsy moth defoliation affects the chemistry of primary host plants … and gypsy moth outbreaks are associated with high densities of pathogens, predators, andparasitoids. At least some of these natural enemies … are known to use alternative hosts. http://www.ars.usda.gov/is/graphics/photos/k7659-1.jpg
Redman & Scriber - OECOLOGIA 125: (2) 218-228 OCT 2000 P. canadensis larvae grew significantly more slowly on leaves from resistant trees, regrowth leaves, and partially eaten leaves from gypsy moth-defoliated areas, than did larvae fed leaves from undefoliated areas All defoliation treatments severely reduced survival … we conducted a long-term growth trial to determine if defoliation by gypsy moths decreased the quality of quaking aspen leaves for P. canadensis. This assay consisted of 40 P. canadensis neonates on each of four treatments: (1) leaves from undefoliated stands, (2) leaves from resistant trees in defoliated stands, (3) regrowth leaves from defoliated stands, and (4) partially eaten leaves from defoliated stands. ("Resistant trees" … had suffered no discernible gypsy moth damage … although they were surrounded entirely by defoliated trees)
Redman & Scriber - OECOLOGIA 125: (2) 218-228 OCT 2000 … increased survival when leaves from defoliated areas were sterilized, whereas sterilization of leaves from undefoliated stands had no effect on survival. … we tested for the presence of pathogenic agents associated with gypsy moth-damaged foliage. P. canadensis neonates (n=25) were placed on the following treatments: (1) leaves from undefoliated stands, (2) surface-sterilized leaves from undefoliated stands, (3) regrowth leaves from defoliated stands, and (4) surface-sterilized regrowth leaves from defoliated stands.
Redman & Scriber - OECOLOGIA 125: (2) 218-228 OCT 2000 Leaves painted with gypsy moth body fluids and water resulted in 100% mortality of swallowtail caterpillars … However, when we autoclaved the body fluid solution, the effect on survival nearly disappeared. We also tested for the presence of pathogenic substances in gypsy moth body fluids themselves. … we collected the cadavers of gypsy moth larvae from an intensely defoliated quaking aspen stand … We ground the cadavers with a mortar and pestle, diluted them with distilled water and painted approximately 1 ml of the resulting solution on quaking aspen leaves
Redman & Scriber - OECOLOGIA 125: (2) 218-228 OCT 2000 When we placed swallowtail caterpillars on trees in areas that had been artificially infested with gypsy moths … the incidence of parasitism increased significantly with the presence of gypsy moths. … we set up a field experiment to test whether gypsy moth larvae attracted parasitoids that attacked P. canadensis larvae as well. Swallowtails were subjected to two treatments: quaking aspen stands artificially infested with gypsy moth larvae, and paired uninfested control stands.
Redman & Scriber - OECOLOGIA 125: (2) 218-228 OCT 2000 Our data suggest that indirect competition between gypsy moths and tiger swallowtails can occur. … gypsy moth-defoliationsignificantly depressed the quality of quaking aspen leaves forPapilio caterpillars, even when leaves were sterilized; thus, leaf chemistry appears to have reduced swallowtail performance. … pathogens or other lethal agents associated with gypsy moth body fluids … could potentially devastate wild tiger swallowtail populations. We observed 84% mortality in a very short period of time on unsterilized leaves …and 100% mortality on leaves painted directly withgypsy moth body fluids. …. as a result of proximity to gypsy moths, parasitism rate was significantly altered. Studies such as this … stand not only to illuminate the basic mechanisms and consequences of interspecific interactions but also to enhance our ability to manage pests responsibly.
Abstract:We examined the nontarget effects of a generalist parasitoid fly, Compsilura concinnata (Diptera: Tachinidae), that has been introduced repeatedly to North America from 1906 to 1986 as a biological control agent against 13 pest species. We tested the effect of previously established populations of this fly on two native, nontarget species of moths (Lepidoptera: Saturniidae), Hyalophora cecropia and Callosamia promethea, in Massachusetts forests. We estimated survivorship curves for newly hatched H. cecropia larvae (n = 500), placed five per tree in the field and found no survival beyond the fifth instar. Figure 1. Survivorship curves for deployed H. cecropia (numbers plotted on a log scale vs. time) recorded from daily observations. Dotted line represents an estimate of the required survival for a population to experience no change in density Effects of a biological control introduction on three nontarget native species of saturniid moths.Boettner et al. 2000. Conservation Biology 14:1798-1806.
Our results suggest that reported declines of silk moth populations in New England may be caused by the importation and introduction of C. concinnata. Boettner et al. We simultaneously deployed cohorts (n = 100) of each of the first three instars to measure the effect of parasitoids during each stage of development. C. concinnata was responsible for 81% of H. cecropia mortality in the first three instars. We deployed semigregarious C. promethea in aggregations of 1-100 larvae in the field and recorded high rates of parasitism by C. concinnata among C. prometheca larvae exposed for 6 days (69.8%) and 8 days (65.6%). We discovered a wild population of a third species of silk moth, the state-listed (threatened) saturniid hemileuca maia maia, and found that C. concinnata was responsible for 36% (n = 50) mortality in the third instar.
The supply of limiting resources, such as nutrients, controls primary productivity; that is, the rate of production of organic matter. On local scales, productivity and diversity are often unimodally related (Fig. 1a), such that peak diversity is observed at intermediate productivity8. Declining diversity at higher levels of productivity is thought to be due to competitive exclusion. Exclusion can be prevented by periodic mortality events, caused by consumers or physical disturbance. These factors also show unimodal relationships (Fig. 1b). In an attempt to unify these patterns theoretically, one study explored how traditional Lotka–Volterra competition models respond to increases in productivity and disturbance frequency9. The study predicted that the effects of disturbance on diversity depend strongly on productivity, and vice versa (for details see Fig. 1c). Physical disturbance and consumer pressure were predicted to give similar patterns. Consumer versus resource control of species diversity and ecosystem functioning. Worm et al. 2002. NATURE 417: 848-851.The most striking feature of life on Earth is its diversity. Consequently, the most fundamental question in ecology is which factors maintain diversity in ecological communities. … we analyse the combined impacts of consumers and nutrient resources on plant diversity.
Worm et al. 2002 We tested these models in a food-web context by experimentally manipulating consumer pressure (absent, present) and nutrient supply (no, low, medium, high nutrient enrichment) in two wave-sheltered rocky shore communities. We chose the two sites to test for consumer–nutrient interactions under contrasting conditions of background nutrient supply and productivity (Open bars = Bald Rock = low; filled bars = Maasholm = high). In Bald Rock {low productivity; open bars}, nutrient enrichment increased diversity and consumers decreased diversity. consumers had strong negative effects under ambient conditions, but weak effects under enriched conditions. As predicted, changes in nutrient supply and consumer pressure had interactive effects on species diversity. In Maasholm {high productivity; filled bars}, the reverse applied: nutrient enrichmentdecreased diversity and consumersincreased diversity. … these effects were interactive: consumers reduced diversity under ambient conditions, but enhanced it under enriched conditions. Peak diversity was found in treatmentswithout consumers in Bald Rock {low productivity}, but in treatments with consumers in Maasholm {high productivity}.
Worm et al. 2002 The effects of nutrient enrichment depend on consumer pressure and vice versa. Both factors have strong and opposing effects on diversity, which change in sign among low-productivity and high-productivity ecosystems. When consumers are present, peak diversity shifts towards higher levels of nutrient supply. These results have important implications for conservation of biodiversity and environmental management because they strongly suggest the potential for synergistic interactions among the most common human impacts on ecosystems. Human alterations of the nitrogen and phosphorus cycles continue to increase nutrient supply and productivity in ecosystems worldwide. At the same time, consumer pressure is altered through overharvesting of herbivore and predator populations, habitat fragmentation and destruction. We conclude that it is not meaningful to assess or manage these impacts in isolation. Rapid change in species composition and loss of diversity will occur when the dynamic balance of consumer and resource control is distorted, especially when consumer removals and resource enrichment occur at the same time.




















