Download
1 / 41
530 likes | 977 Views
( m)RNA Processing. Introduction 1- mRNA capping at the 5’ end ( Euk ) A-Cap structure B-Cap addition C-Significance 2-mRNA Polyadenylation at the 3’ end A- polyadenylation sites B-The polyadenylation machinery C-The poly(A ) polymerase 3-mRNA Splicing

E N D
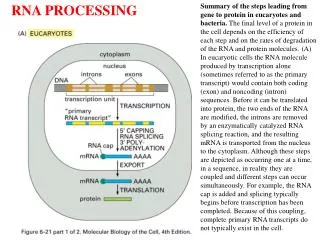
(m)RNA Processing Introduction 1-mRNA capping at the 5’ end (Euk) A-Cap structure B-Cap addition C-Significance 2-mRNA Polyadenylation at the 3’ end A- polyadenylation sites B-The polyadenylation machinery C-The poly(A) polymerase 3-mRNA Splicing A-Splicing sites and signals/mechanism B-Spliceosome assembly and catalysis C-RNA conformational changes 4-Significance of mRNA splicing generating genetic diversity 5-Ribozymes -Group I introns -Group II introns -riboswitches 6- mRNA Degradation Not treated: ribosomal RNA processingMost stable RNAs processing
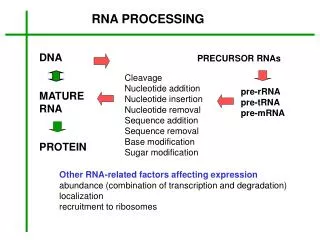
A mature Eukaryotic mRNA After Processing stop AUG m7GpppN1 AAAAAAAA Poly(A) tail CAP Structure Red elements (Cap, polyA tail) are not encoded within the genes: they are added after transcription/co-transcriptionally
The 5’ Cap structure of eukaryotic mRNAs Unusual 5’ppp5’ linkage !!! Presence of methyl Groups at positions 1 and 2 is variable
Why a cap structure ? - Protects mRNA against degradation by 5’->3’ exonucleases • Enhances • translation • initiation by binding of eIF4E
Enzymology of 5’ Cap addition
Why a poly(A) tail ? • Protects mRNA against degradation • by 3’->5’ exonucleases (AAA)n PABI - Facilitates mRNA export out of the nucleus - Facilitates Translation : PAB is a component of the translation machinery
Why a poly(A) tail ? • Enhances • translation • initiation • by binding • of PAB
1st step = mRNA cleavage CFs = Other Cleavage Factors, required for cleavage PAP = Poly(A) Polymerase polymerizes poly (A) tail PAP CFs CPSF CStF AAUAAA CStF = Cleavage Stimulatory Factor 3/4 subunits recognizes the G/U rich element CPSF Cleavage and Polyadenylation Specificity Factor 4 subunits, recognizes the conserved hexamerAAUAAA CPSF 73kD subunit likely responsible for endonuclease activity
The active site of human CPSF-73 suggests an endonuclease activity Zn binding in human CPSF-73 PDB ID= 2I7V Mandel et al.- Nature 2006 Zn binding in L1 metallo b-lactamase
PABII 2nd step = Poly(A) addition ATP CPSF PAP A A3’OH PPi A A A AAUAAA A A A A A A A A A A A A A A A A A A • PABII = Poly(A) Binding Protein II • binds the nascent poly(A) tail when the size is about 10nt • --> provides processivity to poly(A) polymerase • - controls the length of the poly(A) tail (70nt in yeast, 200 in mammals) PAP = chemistry similar to RNA Polymerases…but no template
Structure of the Ternary complex: Poly(A) Polymerase,RNA and ATP Enzyme is a D154A mutant version which allows the crystallization of the ternary complex without reacting From Balbo & Bohm Structure 2007 PDB ID = 2Q66
! An exon does not always correspond to a coding sequence ! An intron does not always correspond to a non-coding sequence ! pre-mRNA Splicing Definitions Intron = RNA piece between 2 exons that is excised during splicing Exon = RNA sequence that is spliced with another exon First proof of the existence of “split” genes: Hybridization of a gene DNA with the corresponding mature mRNA shows that they are not colinear intron exon
- 1 2 3 4 Intron-exon2 Intron Precursor Ligated exons + Exon/ introns junctions and consensus sequences 30nt 10nt-several kb 10-30nt Exon2 Exon1 GUAUGU UACUAAC Poly(Y) YAG Branchpoint 3’ splice site 5’ splice site Poly Pyrimidine Tract In vitro pre-mRNA splicing reaction: • synthesis of an RNA substrate containing exon1, intron , exon2 and splicing signals • Incubation with nuclear extract with MgCl2 and ATP 1 - Extract, MgCl2 No ATP 2- Extract, ATP and MgCl2 3 - RNA substrate with a 5’ splice site mutation 4 - RNA substrate with a 3’ splice site mutation • Deproteinization, RNA extraction and fractionation of products by denaturing gel electrophoresis • Visualization of products by autoradiography
Lariat Intron Ligated Exons (5’-3’phosphodiester) Lariat Intermediate Each chemical step is a transesterification 1st step: formation of a 2’-5’ phosphodiester link 2nd step: formation of a 5’-3’ phosphodiester link The chemical steps per se do not require energy The ATP is required for steps other than the chemical steps
Pre-mRNA splicing is catalyzed by a complex machinery: the Spliceosome 145 spliceosomal proteins 45nm snRNPs: small nuclear ribonucleoprotein particles The U1 snRNP Each snRNP contains several proteins+ 1 small nuclear RNA U1, U2, U4, U5, U6
Recognition of Splicing Signals by base-pairing with the snRNAs of snRNPs U1 snRNA base pairs with the 5’ splice site U2 snRNA base pairs with the branchpoint
U1 CCAUUCAUA1-5’ UGGUAUGUUC 5’- Splicing U1 CCCUUCAUA1-5’ UGGGAUGUUC 5’- Splicing (of this particular substrate) is restored by a compensatory mutation in U1 How do we demonstrate experimentally these base pairing interactions ? U1 CCAUUCAUA1-5’ UGGGAUGUUC 5’- No Splicing U1 CCGUUCAUA1-5’ UGGGAUGUUC 5’- Suppression is specific: A->G mutation in U1 does not restore splicing
How is the branchpoint recognized ? base pairing with the U2 snRNA confirmed by compensatory mutations The branchpoint must be in a “bulged” conformation (unpaired and extruded from the duplex) to be selected as the nucleophile
BBP recognizes the branchpoint Adenosine prior to U2 binding Multiple steps of branchpoint recognition PDB ID = 1K1G StaleyNSB 2002
Early recognition of splice sites by the spliceosome 5’exon GUAUGU U1 snRNP 3’exon CAAUCAU YYYYY BBP/SF1 Mud2/U2AF 1st complex to assemble specifically on the pre-mRNA No ATP is required recognizes both the 5’SS and the branchpoint bridges the 5’ and the 3’ splice sites
Spliceosome Assembly
AG GU U6 A U1 AG U2 GU U6 U1,U4 U6 U6 A intron U2 Exons U4 snRNAs RNA Conformational changes prior to catalysis •U6 replaces U1 for recognition of the 5’ splice site •U4 is replaced by U2 for base pairing to U6: thought to promote formation of a catalytically active structure • RNA remodeling requires ATP-dependent RNA helicases and ATP hydrolysis
A metal binding site required for splicing catalysis in the U6 snRNA Demonstrated by Metal Specificity Switch experiments • Phosphate Oxygens (“hard” Lewis base) in D/RNA often coordinate Mg++(“hard” Lewis acid) • Sulfur substitution of these oxygens(phosphorothioate) block Mg++ binding because it is a softer Lewis base than O in phosphates Mn++ or Cd++ can bind S because they are soft Lewis acids; if the phosphate serves as a divalent cation binding site, Mn/Cdcan sometimes rescue the defect introduced by a phosphorothioate Sontheimer NSB 2001 8, 11-13
Rescue of the splicing defect induced by a phosphorothioate substitution of U6-U80(Sp) by addition of Mn2+ or Cd2+ Koodathingal et al. Molecular Cell 2010
A two metal ion mechanism for the splicing reaction (remember T.Steitz and DNA/RNA/PolyA pol.) J.Steitz T.Steitz
Generating Diversity through alternative splicing Architecture of the Dscam gene, an extracellular receptor required for axon guidance in Drosophila Exon 4: 12 possible exons sequences Exon 6: 48 possible exons sequences Exon 9: 33 possible exons sequences Exon 17: 2 possible exons sequences Exons 4, 6, 9 and 17 have multiple possible sequences that are chosen through alternative splicing Total possible combinations =12 X 48 X 33 X 2= 38,016 Number of genes in the Drosophila genome = 20,000-50,000….
How do cells choose which exons to splice ? • Exons and introns possess regulatory sequences onto which splicing regulatory proteins bind • Splicing regulators (SR proteins or others) have affinity for specific enhancers or silencers (same as for transcription factors). • Splicing regulators (SR proteins or others) promote or repress recruitment of spliceosomes at nearby splice sites resulting in inclusion or skipping of exons Enhancer X Enhancer Y Enhancer Z Intronic repressor Intronic enhancer
mRNA Splicing can also produce functional RNA molecules from introns: Exons are not necessarily expressed and introns are not necessarily useless sequences:The example of the UHG gene Exon1 Exon2 Exon3 small RNA1 small RNA2 intron2 intron1 Splicing Exons are degraded and not translated (no ORF) Lariat debranching and Further processing Mature small RNAs Many small RNAs such as microRNAs are contained in intronic sequences
Study of other type of splicing systems reveals the existence of Ribozymes = RNA molecule capable of catalyzing a (bio)chemical reaction Examples of natural ribozymes Namereaction (Genomic) localization Group I self-splicing Nuclear rDNATetrahymena Introns organelles of fungi and plants bacteriophages, bacteria Group II self-splicing organelles fungi and plants introns splicing pathway bacteriophages is identical to bacteria nuclear pre-mRNA introns Non-Splicing Ribozymes: RNase P 5’ end cleavage Eukaryotes of pre-tRNAs Prokaryotes RibosomePeptidyl Eukaryotes Transferase Prokaryotes T.Cech Chemistry Nobel Prize 1989 S.Altman
Group I introns self-splicing Group II introns self-splicing •No exogenous nucleophile: an internal adenosine is the nucleophile for the 1st step •The splicing pathway is identical to the pathway used to excise spliceosomal introns An exogenous nucleophile, a Guanosine nucleoside is the nucleophile for the 1st step
IV Tertiary Structure Ribozyme activity Self-splicing in the absence of proteins or snRNAs The two splicing steps are identical to those of nuclear splicing Autocatalytic group II introns suggest that pre-mRNA splicing might be RNA-catalyzed Similarities between group II introns domain5 and the U6 stem-loop group II introns secondary structure binds Mg2+ binds Mg2+ +monovalentcations +Mg++ C GA Biochemical Society Transactions (2005)
Group II introns domain 5 binds two divalent cations involved in catalysis -> Suggests that these follow the 2-metal ion catalysis model proposed by Steitz and Steitz 358 377 PDB ID = 3EOH Toor et al. Science. 2008 Apr 4;320(5872):77-82.
w Self-splicing of the Tetrahymena Pre-rRNA group I Ribozyme G/u 3’ GOH cucucu/A 5’ Pre-rRNA + GOH Intronfolding; Binding of Guanosinein the G binding site GGGAGG w G/u 3’ First Transesterification cucucu-OH 5’ GGGAGG AG5’ G/u Conformational Change: Binding of the wG In the G binding site 3’ w cucucu-OH 5’ GGGAGG AG5’ Excised Intron +guanosine at the 5’end Second Transesterification G-OH Red=intron sequences Blue=exon sequences Green= G cofactor GGGAGG cucucu/u 3’ 5’ AG5’ Ligatedexons
Group I Ribozymes catalyze RNA reactions by forming a Complex 3D structure -like protein enzymes- PDB ID = 1ZZN • Monovalent and Divalent Cations required to prevent electrostatic repulsion between strands (Purple Sphere = K+) (Green Spheres = Mg++) Tertiary interactions help fold the RNA into 3D structure: • Pseudoknots • tetraloop-tetraloop Receptors interactions • Ribose zippers • A-minor interactions Stahley and StrobelScience Vol. 309. no. 5740, pp. 1587 - 1590
Tertiary interactions that help fold the RNAs into complex 3D structures: pseudoknot/kissing loops tetraloop/receptor ΔG: −2 to −4 kcal mol.−1 ΔG: −6 to −15 kcal mol.−1 Ribose zippers A-minor interaction ΔG: less than −1 kcal mol.−1 ΔG: less than −1 kcal mol.−1
A model of the first step Active Site of Group I ribozymes Exon1 O P O- O O G C O OH O P O- O O G U Metal binding site O G-binding site =G130-C177 Intron Mg++ O OH G130 O The G-binding site binds both the guanosine cofactor and the wG during the successive steps ->structural evidence for a single active site P O N9 H O N7 N3 OH O H N 2’ H 3’ N N3 N1 O6 1’ H N1 (The predicted transition state is shown) H 5’ H O N9 O H OH O N7 N3 N Guanosine cofactor H N1 C177
Where is the Metal binding site ? A published structure provides a clue (for the 2nd step) Catalysis Model PDB ID = 1ZZN Crystal Structure Stahley and StrobelScience Vol. 309. no. 5740, pp. 1587 - 1590
Ribozymes can be used to Control of Gene Expression:example of a metabolite-responsive ribozyme glmS: catalyzes the production of Glucosamine N6-phosphate (a precursor of cell wall components) CechNature 2004
Control of mRNA Degradation Elements that need to be removed for efficient mRNA degradation: The cap structure will allow access to exoribonucleases for digestion The poly(A) tail Major pathway in eukaryotic cells: Deadenylation Decapping Exonuclease Digestion of the body of the mRNA Deadenylases Decapping complex (Dcp) 5’->3’ Exonucleases: Rat1 (nucleus,) Xrn1 (Cytoplasm) 3’->5’ Exonucleases: The Exosome (N & C)
Functional Compartmentalization of exonucleases activities in eukaryotic cells Rrp6p
RNAi-mediated degradation of mRNAs: Natural and Experimental ways to control mRNA levels in eukaryotic cells Medicine Nobel Prize 2006 Slicing = Cleavage in the body of the mRNA provides entry sites for 5’-3’ and 3’-5’ exonucleases and degradation