Download

1 / 63
670 likes | 992 Views
Hormone Receptors on the Plasma Membrane. Characteristics of Receptors in General Five Groups of Membrane-Bound Receptors The G Protein-Coupled Receptor Superfamily Signal Transduction through Cyclic AMP Signal Transduction through Phospholipase C Role of Calcium Role of Protein Kinase C.

E N D
Hormone Receptors on the Plasma Membrane Characteristics of Receptors in General Five Groups of Membrane-Bound Receptors The G Protein-Coupled Receptor Superfamily Signal Transduction through Cyclic AMP Signal Transduction through Phospholipase C Role of Calcium Role of Protein Kinase C
General Characteristics of Receptors • Receptors bind hormones, resulting in a biological response • All receptors exhibit general characteristics: - Specific Binding (structural and steric specificity) - High Affinity (at physiological concentrations) - Saturation (limited, finite # of binding sites) - Signal Transduction (early chem event must occur) - Cell Specificity (in accordance with target organ specificity).
LH hCG FSH FSH hCG LH LH Receptors Specific Binding • A receptor will only bind (recognize) a certain hormone, or closely related hormones.
Receptors Have High Affinity • In the bloodstream, there are thousands of different peptides. • Hormones are present in very small quantities (nanogram or picogram). • Receptors must therefore be very sensitive to the presence of a hormone (they must be able to bind the hormone even if it is present in low amounts). • Thus, they have high affinity (ability to bind at low hormone concentrations).
Analysis of Receptor Binding Sites TESTIS 125 Na- I + hCG lactoperoxidase method 125 I -hCG TRACER RT O/N WASH, PBS Count Pellet CPM Centrifuge Leydig/Interstitial Cells Seminiferous Tubules
Criteria for hormone-mediated events • Receptor must possessstructural and steric specificity for a hormone and for its close analogs as well. • Receptors are saturable and limited (i.e. there is a finite number of binding sites). • Hormone-receptor binding is cell specific in accordance with target organ specificity. • Receptor must possess ahigh affinity for the hormone at physiological concentrations. • Once a hormone binds to the receptor, some recognizable early chemical event must occur.
Affinity: The tenacity by which a drug binds to its receptor. • Discussion: a very lipid soluble drug may have irreversible effects; is this high-affinity or merely a non-specific effect? • Intrinsic activity: Relative maximal effect of a drug in a particular tissue preparation when compared to the natural, endogenous ligand. • Full agonist – IA = 1 (*equal to the endogenous ligand) • Antagonist – IA = 0 • Partial agonist – IA = 0~1 (*produces less than the maximal response, but with maximal binding to receptors.) • Intrinsic efficacy: a drugs ability to bind a receptor and elicit a functional response • A measure of the formation of a drug-receptor complex. • Potency: ability of a drug to cause a measured functional change.
Receptors have two major properties: Recognition and Transduction Recognition: The receptor protein must exist in a conformational state that allows for recognition and binding of a compound and must satisfy the following criteria: • Saturability – receptors exists in finite numbers. • Reversibility – binding must occur non-covalently due to weak intermolecular forces (H-bonding, van der Waal forces). • Stereoselectivity – receptors should recognize only one of the naturally occurring optical isomers (+ or -, d or l, or S or R). • Agonist specificity – structurally related drugs should bind well, while physically dissimilar compounds should bind poorly. • Tissue specificity – binding should occur in tissues known to be sensitive to the endogenous ligand. Binding should occur at physiologically relevant concentrations.
The failure of a drug to satisfy any of these conditions indicates non-specific binding to proteins or phospholipids in places like blood or plasma membrane components.
Receptors have two major properties: Recognition and Transduction Transduction: The second property of a receptor is that the binding of an agonist must be transduced into some kind of functional response (biological or physiological). Different receptor types are linked to effector systems either directly or through simple or more-complex intermediate signal amplification systems. Some examples are: • Ligand-gated ion channels – nicotinic Ach receptors • Single-transmembrane receptors – RTKs like insulin or EGF receptors • 7-transmembrane GPCRs – opioid receptors • Soluble steroid hormones – estrogen receptor
Predicting whether a drug will cause a response in a particular tissue Factors involving the equilibrium of a drug at a receptor. • Limited diffusion • Metabolism • Entrapment in proteins, fat, or blood. Response depends of what the receptor is connected to. • Effector type • Need for any allosteric co-factors – THB on tyrosine hydroxylase. • Direct receptor modification – phosphorylation
Receptor theory and receptor binding. Must obey the Law of Mass Action and follow basic laws of thermodynamics. • Primary assumption – a single ligand is binding to a homogeneous population of receptors NH+3 COO-
kon/k1 [ligand] + [receptor] [ligand receptor] koff/k2 • kon = # of binding events/time (Rate of association) = [ligand] [receptor] kon = M-1 min-1 • koff = # of dissociation events/time (Rate of dissociation) = [ligand receptor] koff = min-1 • Binding occurs when ligand and receptor collide with the proper orientation and energy. • Interaction is reversible. • Rate of formation [L] + [R] or dissociation [LR] depends solely on the number of receptors, the concentration of ligand, and the rate constants kon and koff.
At equilibrium, the rate of formation equals that of dissociation so that: [L] [R] kon = [LR] koff KD = k2/k1 = [L][R] [LR] *this ratio is the equilibrium dissociation constant or KD. KD is expressed in molar units (M/L) and expresses the affinity of a drug for a particular receptor. • KD is an inverse measure of receptor affinity. • KD = [L] which produces 50% receptor occupancy
Once bound, ligand and receptor remain bound for a random time interval. • The probability of dissociation is the same at any point after association. • Once dissociated, ligand and receptor should be unchanged. • If either is physically modified, the law of mass action does not apply (receptor phosphorylation) • Ligands should be recyclable.
Receptor occupancy, activation of target cell responses, kinetics of binding • Activation of membrane receptors and target cell responses is proportional to the degree of receptor occupancy. • However, the hormone concentration at which half of the receptorsis occupied by a ligand (Kd) is often lower than the concentration required to elicit a half-maximal biological response(ED50)
Receptor Fractional Occupancy F.O. = [LR]____ = [LR]___ *now substitute the KD equation. [Total Receptor] [Rf] + [LR] [R] = KD • [LR] F.O. = [Ligand] [L] [Ligand] + KD Use the following numbers: [L] = KD= 50% F.O. [L] = 0.5 KD = 30% F.O. [L] = 10x KD = 90%+ F.O. [L] = 0= 0% F.O. 100 Fractional Occupancy 50 0 Ligand Concentration
Assumptions of the law of mass action. • All receptors are equally accessible to ligand. • No partial binding occurs; receptors are either free of ligand or bound with ligand. • Ligand is nor altered by binding • Binding is reversible • Different affinity states?????
Studies of receptor number and function • We can directly measure the number (or density) of receptors in the LR complex. • Ligand is radiolabeled (125I, 35S. or 3H). Selection of proper radioligand: • Agonist vs. antagonist (sodium insensitive) • Higher affinity for antagonists • Longer to steady state binding • Saturation binding curve-occurs at steady state conditions (equilibrium is theoretical only). • Demonstrates the importance of saturability for any selective ligand. • Provides information on receptor density and ligand affinity and selectivity.
Scatchard transformation • Y-axis is Bound/Free (total radioligand-bound) • X-axis Bound (pmol/mg protein) • Straight lines are easier to interpret.
The amount of drug bound at any time is solely determined by: • the number of receptors • the concentration of ligand added • the affinity of the drug for its receptor. • Binding of drug to receptor is essentially the same as drug to enzyme as defined by the Michelis-Menten Equation.
100 % binding 50 Kd 0 10 30 100 300 1000 Dose of Hormone Thus, to reiterate…,Calculating Affinity • Take a cell which has the receptor on it (ie, granulosa cells with FSH receptor). • Prepare membrane homogenate. • Incubate membranes with increasing amounts of labeled hormone. • Determine how much binding of hormone occurs at each dose. • Dissociation constant (Kd) is dose where 50% of maximal binding occurs.
100 saturation % binding 50 0 10 30 100 300 1000 Dose of Hormone Thus, to reiterate…,Saturation • There is a finite limit to the numbers of receptors which can be on a cell. • Therefore, there’s a maximum amount of binding which can occur (all receptors are saturated)
Biological Response to Ligand Binding • A receptor not only binds hormone; there must also be a biological response from the cell (e.g., increased transcription, phosphorylation, etc.) • This is also called “signal transduction”. • The biological response can result from: - the ligand itself (e.g., Fe, LDL) - the receptor (e.g., increased cyclic AMP, transcription, phosphorylation)
Determinants of Biological Response • The strength of the response of the cell to the hormone depends upon three factors: 1) the amount of hormone present to bind to the receptors 2) the numbers of receptors on the cell 3) the affinity of the receptor for the hormone (how much hormone do you need to get receptor binding?)
100 50 0 Biological Response (% max) 0 25 50 75 100 Receptor Occupancy (%) Regulation of Receptor Number:the Phenomenon of Spare Receptors • We know that cells typically have about 20 times more receptors than is needed for a maximal biological response. • A complete biological response occurs after binding to only 5% of the receptors on a cell. • This remaining 95% are called “spare receptors”. • Why have spare receptors?
100 75 50 25 0 100 75 50 25 0 % maximal response % of receptors occupied Kd 10-11 10-10 10-9 Hormone Concentration (M) Effect of Decreasing Receptor Number in a Cell Which Does Not Have Spare Receptors • No change in affinity. • Decrease in maximal biological response.
0 50 75 20,000 15,000 10,000 5,000 0 Number of receptors bound % Reduction in Receptor Number # Receptors for maximal biological response Kd Hormone Concentration (M) Effect of Decreasing Receptor Number in a Cell Which Has Spare Receptors • In this example, assume you need 5000 receptors occupied for maximal biological response. • If you start w/ 20,000 receptors occupied, decreasing receptor number does not change receptor affinity (Kd).
10,000 100 80 60 40 20 0 5000 20,000 R/cell % Maximal Biological Response 2500 10-11 10-10 10-9 10-8 10-7 Hormone Concentration (M) Effect of Decreasing Receptor Number in a Cell Which Has Spare Receptors • No change in maximal biological response (unless you go below 5000 receptors/cell). • Requires higher dose of hormone to obtain maximal response.
Hormones, Agonists, and Antagonists • Substances other than a receptors normal hormone may exist (or be made). • Each substance that binds to a receptor has an intrinsic activity (related to the resulting biological response). Substance Intrinsic Activity hormone 100% (by definition) superagonist >100% partial agonist <100% antagonist 0%
progesterone RU486 hormone binding domain transcriptional domain PR Antagonists • If a substance binds to a receptor but does not cause a biological response, it blocks the natural hormone from binding to it. • Example: RU486 binds to the progesterone receptor, but does not cause a response. DNA
Regulation of Biological Response at the Receptor Level • The strength of signal transduction can be regulated at the level of the receptor by several mechanisms: 1) change the affinity of the receptor (make it bind more difficult or easier to bind hormone). This usually doesn’t happen. 2) change the numbers of receptors on the cell (common) - internalization and degradation of receptors - occupancy of receptors (prevents hormone binding) - gene expression/synthesis 3) change the signal transducing ability of the receptor (usually for rapid regulation) - phosphorylation (usually inhibits receptor activity) - G protein uncoupling (stay tuned)
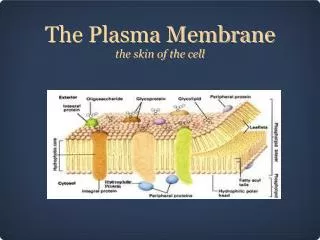
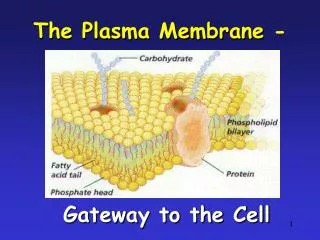
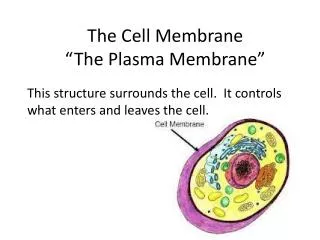
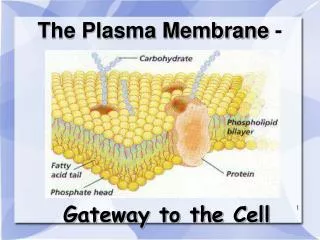
Plasma Membrane-Bound Receptors • Recall that peptide hormones are polar, and cannot readily cross the cell membrane. • Therefore, their receptors must be on the outside surface of the cell.
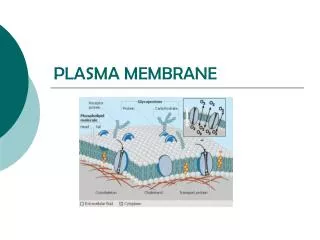
Types of Plasma Membrane Receptors There are five basic types of membrane bound receptors (grouped by signal transduction method): • tyrosine kinase receptors • receptors that are closely linked to tyrosine kinases • receptors with guanylyl cyclase activity • receptors that serve as transporters • G protein-coupled receptors
extracellular domains (ligand binding) plasma membrane tyrosine phosphorylase domains phosphorylated enzyme (altered activity) Signal Transduction by Plasma Membrane Receptors 1) Receptors with intrinsic tyrosine kinase activity. Binding of hormone to the receptor induces the phosphorylating activity of the receptor. Example: Insulin receptor
associated tyrosine kinase phosphorylated enzyme Signal Transduction by Plasma Membrane Receptors 2) Receptors that are closely linked to tyrosine kinases. These activate cytoplasmic tyrosine kinase enzymes. Example: Growth Hormone Receptor
guanylate cyclase GTP ion channels cyclic GMP protein kinase G phosphodiesterase levels Signal Transduction by Plasma Membrane Receptors 3) Receptors with Guanylyl Cyclase Activity. Binding to the receptor activates guanylate cyclase region of the receptor, causing conversion of GTP to cyclic GMP. Example: Atrial Natriuretic Peptide Receptor
iron transferrin Signal Transduction by Plasma Membrane Receptors 4) Receptors that serve as transporters. These move the ligand inside the cell, where they have an effect. (Not typical for hormones). Example: Iron, transported by transferrin receptor
Signal Transduction by Plasma Membrane Receptors 5) G Protein-coupled receptors. (The largest group!) These receptors are coupled with guanine nucleotide-binding proteins (G proteins), which activate various signaling pathways. Examples: Receptors for LH, FSH, TSH, GnRH, dopamine, serotonin, glutamine, parathyroid hormone, interleukins, etc.
-NH 2 COOH- G Protein-Coupled Receptor Superfamily • Common structural features: - an amino terminus hormone-binding domain - seven hydrophobic transmembrane domains - a carboxyl terminus, intracellular domain
G Protein-Coupled Receptor Superfamily • Common functional features: - binding to the receptor activates a G protein - each receptor is associated with a specific type of G protein - each G protein type has different functions: - Gs: stimulates cyclic AMP - Gi: inhibits cyclic AMP - Go activates phospholipase C
NH2 g b a COOH GDP How G Proteins Work • G proteins are composed of three subunits: alpha (a), beta (b), and gamma (g). • In the inactive state, the three subunits are associated with the receptor at the plasma membrane. The alpha subunit has a guanosine diphosphate attached.
hormone hormone NH2 NH2 g g b b a COOH COOH a GDP GTP GTP signal pathways How G Proteins Work (cont.) • When hormone binds, the GDP leaves the alpha subunit, and is replaced by a GTP. • The alpha subunit then goes off to activate signaling pathways.
NH2 g b a COOH a GDP GTP P How G Proteins Work (cont.) • After activating the signal pathway, the GTP is hydrolyzed into GDP, and the alpha subunit returns to the beta and gamma subunits at the membrane.
G Protein Stimulation of Cyclic AMP • Binding of many hormones to their receptors results in the stimulation of the second messenger, cyclic AMP. • G Protein involved: Gs
LH a a Gs b b g LHR AC b a GDP GTP a GTP b ATP GTPase cAMP GTP GDP G Protein Stimulation of Cyclic AMP
Receptor-G protein InteractionsHow are receptor-G protein interactions measured? Low- affinity R + G(GTP-δ-S) RG(GDP) GTPγS GDP Without GTP, both high- and low-affinity states are measured. With GTP and Mg2+, only low-affinity state is measured, because Agonist binding rapidly induces change from high- to low-affinity. Ligand-binding assays: High-affinity
How Else Is Cyclic AMP Regulated? • In addition to regulating the production of cyclic AMP, there is also regulation of its degradation. • Degradation of cyclic AMP is by phosphodiesterases, which break down cyclic AMP into 5’-AMP • Inhibitors of phosphodiesterases prolong activation of the cyclic AMP system.
a b ATP cAMP GTP (-) PDEs 5’-AMP Regulation of cAMP Levels by Phosphodiesterases X PDE Inhibitor