Download

1 / 56
600 likes | 893 Views
DNS, RNS un proteīnu biosintēze. Trīs galvenie procesi šūnā. Replikācija (DNS-DNS) Transkripcija (DNS-RNS) Translācija (RNS-proteīni). Kurš process ir visnepieciešamākais dzīvībai?. Translācija ? Transkripcija ? Replikācija ?. DNS. RNS. Proteīni. Informācijas plūsma.

E N D
Trīs galvenie procesi šūnā • Replikācija (DNS-DNS) • Transkripcija (DNS-RNS) • Translācija (RNS-proteīni)
Kurš process ir visnepieciešamākais dzīvībai? • Translācija ? • Transkripcija ? • Replikācija ? DNS RNS Proteīni Informācijas plūsma Informācijas nesēja replikācija
Galvenās atšķirības starp eikariotu un prokariotu šūnām • Eikariotos kodolam un citām organellām ir sava membrāna • Prokariotiem nav citoskeleta • Genoma organizācija ir ļoti atšķirīga • Vērojamas lielas atšķirības bioķīmiskajos procesos, replikācijā, transkripcijā un translācijā
Kas notiek replikācijā? 1. DNS dubultspirāle atritinās 2. Pēc komplementaritātes principa uz abiem esošajiem DNS pavedieniem veidojas jauni
OH OH OH OH OH OH OH O O O O O O O OH OH OH O P O P OH OH OH O 5'gals Replikācijas ķīmija P P CH2 CH2 Base Base O O P P CH2 CH2 Base Base O O H20 + P 3' P OH P + Saites veidošanās CH2 P Base O CH2 5' Base O OH 3' 3'gals OH
Kas ir nepieciešams replikācijai ? • 1) dsDNS • 2) Oriģins – DNS rajons, kuru atpazīst replikācijas mašinērija • 3) Replicējošie enzīmi • 4) Nukleotīdi
Polimerāžu enzimātiskās aktivitātes • 5’-3’ polimerāzes aktivitāte 5’-AAGTCACC-3’ 5’-AAGTCACCG-3’ 3’-TTCAGTGGCAA-5’ 3’-TTCAGTGGCAA-5’ NEKAD nav 3’-5’ polimerāzes aktivitāte! +G 5’-3’ polimerāzes aktivitāte piemīt visām DNS un RNS polimerāzēm
Polimerāžu enzimātiskās aktivitātes • 3’-5’ eksonukleāzes (editēšanas) aktivitāte 5’-AAGTCAC -3’ 5’-AAGTCAC-3’ 3’-TTCAGTGGCAA-5’ 3’-TTCAGTGGCAA-5’ A -A • Normālos apstākļos, tiek aizvākts viens nepareizi pievienots nukleotīds (t.i., izlabota kļūda) • 3’-5’ eksonukleāzes aktivitāte piemīt lielākajai daļai (bet ne visām) DNS un RNS polimerāžu • Dažām polimerāzēm piemīt arī 5’-3’ eksonukleāzes aktivitāte (piemēram, DNS polimerāzei I)
Polimerāzes un praimeri • DNS polimerāzes spēj tikai pagarināt esošu DNS vai RNS fragmentu (praimeri), bet nespēj uzsākt fragmenta sintēzi • RNS polimerāzēm ir nepieciešams tikai komplimentārais pavediens • Tādēļ, replikācijas procesu uzsāk RNS polimerāze (t.i., uzsintezē praimeri), bet turpina DNS polimerāze
Praimeris Komplimentārais pavediens (matrica)
DNS replikācija (E.coli) Pol III sintezē vadošo pavedienu 4 2 1 Helikāze atvij spirāli Primāze sintezē RNS praimeri 3 Topoizomerāze sašķeļ vienu DNS pavedienu, lai noņemtu mehānisko spriegumu Pol I aizvieto RNS praimeri ar DNS 5 6 7 SSB proteīni aizkavē dubultspirāles veidošanos Pol III pagarina praimeri, producē Okazaki fragmentu DNS ligāze savieno Okazaki fragmentus
DNS Replikācija (Eikarioti) RPA proteīni aizkavē dubultspirāles veidošanos Pol dsintezē vadošo pavedienu 5 2 1 Helikāze atvij spirāli Primāze sintezē RNS praimeri 3 4 Topizomerāze sašķeļ vienu DNS pavedienu, lai noņemtu mehānisko spriegumu RNāze H izšķeļ RNS praimeri Pol a nedaudz pagarina RNS praimeri 5 6 7 Pol d aizvieto Pol a;producē Okazaki fragmentu DNS ligāze savieno Okazaki fragmentus
Atšķirības eikariotu un prokariotu replikācijas dakšā • Eikariotos RNS praimeri vispirms pagarina Pol α un tad Pol δ. Prokariotos praimeri pagarina tikai PolIII • Eikariotos RNS praimeri izšķeļ RNāze, kamēr prokariotos – DNS PolI, kura vienlaicīgi arī aizpilda spraugu ar DNS • Prokariotos Okazaki fragmenti ir aptuveni 2000 bp gari, bet eikariotos – aptuveni 200bp gari (nevis otrādi...)
Citas atšķirības eikariotu un prokariotu replikācijā • Cēloņi: • 1. Eikariotiskās hromosomas ir daudz garākas par prokariotiskajām • 2. Eikariotiskās hromosomas ir lineāras, nevis cirkulāras • 3. Eikariotiskās hromosomas ir sapakotas un uztītas uz histoniem
Hromosomu multiplie oriģini Eikarioti Baktērijas
DNS sintēzes ātrumi un nepieciešamība pēc vairākiem oriģiniem Oriģini Genoms Repl. ātrums Repl. laiks Komentāri 30 kb/min 40 min 1 4.6 Mbp E. coli Repl.ilgtu 80st ja būtu 1 ori 14 Mbp (1 cm) 3 kb/min 20 min 330 Raugs 1 l kultūrā = 4.1010šūnu --> 400 000 km sintezētas DNS Repl. ilgtu 1 gadu, ja būtu 1 ori 3 Gbp (2 m) 3 kb/min 7 h >10 000 ? Cilvēks Dzīves laikā (1016šūnu dalīšanās)sintezēti 2x1013 km DNA (2 gaismas gadi)
Lineāras DNS replicēšanai vajag īpašus līdzekļus: Telomēras un telomerāze • Telomēras: īsu sekvenču atkārtojumi hromosomu galos • Telomerāze: polimerāze, kas veido telomēras • Kam noder telomēras un telomerāze?
Telomerāze darbībā • Telomerāze satur savu RNS fragmentu, kas darbojas kā matrica DNS galu sintēzei • RNS fragments satur divus tiešos atkārtojumus (parādīts aaaacccc 2x atkārtojums dažos vienšūņos, mugurkaulniekiem tas ir cccctaa) • Pēc viena nukleotīdu pievienošanas cikla, telomerāze translocējas līdz nākošajam ttttgggg atkārtojumam • Realitātē process ir ievērojami komplicētāks...
Vai telomerāze vienmer ir aktīva? • Aktīva bērniem un pieaugušo dzimumšūnās un cilmes šūnās • Neaktīva lielā daļā somatisko šūnu pieaugušajiem • Tātad hromosomas patiešām ar laiku paliek īsākas – viens no iemesliem, kāpēc mēs novecojam • Tā paša iemesla dēļ kultivētas primārās zīdītāju šūnas dalās ierobežotu reižu skaitu • Telomerāzes aktivācija pieaugušās pelēs paildzina to mūžu • Telomerāze ir aktīva lielākajā daļā audzēju
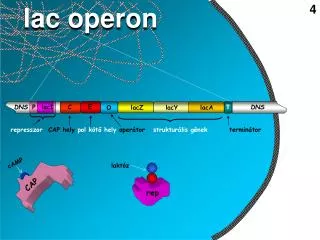
Kas ir nepieciešams transkripcijai? • dsDNS matrica • promoters = transkripcijas uzsākšanas vieta • ribonukleotīdi • RNS polimerāze un transkripcijas faktori
Eksoni, introni un sekvences gēnu galos • Introni –nekodējošās sekvences eikariotu gēnu vidus daļā • Eksoni – gēnu kodējošās sekvences • Introni bieži tiek uzlūkoti kā “nederīga” DNS, bet tie var saturēt regulatoras sekvences vai funkcionālas nekodējošās RNS • Parasti introni ir daudz garāki par eksoniem • Gēnu 5’un 3’ gali satur t.s. netranslētos reģionus (UTR), kuri satur regulējošas sekvences Eksoni 5’ UTR (Untranslated region) 3’UTR Introni
Eikariotiskie promoteriTATA bokss • Vislabāk izpētītā promoteru sekvence • Atrodas daudzu, bet ne visu gēnu 5’ UTR sastāvā 25-35 nukleotīdus pirms transkripcijas sākuma vietas • Piesaista t.s. TATA boksa piesaistīšanās proteīnu TBP, kas ir transkripcijas faktora TFIID sastāvdaļa
Citi eikariotisko promoteru elementi -2 līdz +4 +28 līdz +32 -37 līdz -32 -31 līdz -26 -100 līdz -1 DPE Inr CpG TATA BRE • CpG salas – CG bagātas sekvences, piedalās transkripcijas regulācijā, C nukleotīds var būt metilēts • BRE (TFII Brecognition element) – tieši piesaista transkripcijas faktoru IIB • Iniciatora elements (Inr) darbojas līdzīgi TATA boksam un arī piesaista TBP • DPE elements (Downstream promoter element) piesaista citas TFIID subvienības- TAF9 un TAF5 • Eikariotisko promoteru sastāvā var būt viens vai vairāki no minētajiem elementiem GGG CCA GAC A T T A A G AC TT TATA AA PyPyAN PyPy CGCG G
Enhānseri • Transkripciju regulējošie elementi tālu (līdz 50,000 bp) no transkripcijas sākuma vietas • Var atrasties pirms gēna, pēc gēna vai intronā • Kalpo kā regulatoro proteīnu piesaistes vieta
Vispārējie transkripcijas faktori (GTFi, General transcription factors) • Nepieciešami RNS polimerāzes piesaistei un transkripcijas uzsākšanai no lielākās daļas promoteru • Prokariotos ir tikai viens GTF – sigma faktors • Eikariotos ir vismaz 6 GTFi – TFIIA, TFIIB, TFIID, TFIIE, TFIIF un TFIIH
GTFu funkcijas A B • (A) TFIID (kura sastāvdaļa ir TBP) atpazīst TATA boksu • (B) TFIIA un B stabilizē kompleksu, rada vietu RNS polimerāzes piesaistei • (C) TFIIF piesaista brīvu RNS polimerāzi un kopā ar TFIIE un H piesaistās esošajam kompleksam • TFIIE un TFIIH ir iesaistīti dsDNS pavedienu atdalīšanā • (D) Pēc visu faktoru un RNS polimerāzes piesasitīšanās sākas transkripcija • Pēc transkripcijas uzsākšanas, GTFi no kompleksa disociē C D
Transkripcijas trīs fāzes Start Stop Iniciācija • Iniciācijā pie promotera piesaistas GTFi un RNS polimerāze un izveidojas transkripcijas burbulis • Elongācijā notiek RNS ķēdes pagarināšana • Terminācijā polimerāze sasniedz transkripcijas «stop» sekvenci (terminatoru) un RNS ir gatava • movie Transkripcijas burbulis Elongācija DNS-RNS hibrīds Augošā RNS Terminācija
Pēc transkripcijas... • Prokariotos RNS ir gatava translācijai uzreiz pēc transkripcijas vai pat vienlaicīgi ar to • Eikariotos vispirms ir jāveic RNS procesēšana un transports uz citoplazmu
pre-mRNS procesēšana (1) • 1) Kepings • 2) Poliadenilēšana • 3) Splasings (2) (3)
Keps (angl. cap) 7-metil guanilāts • Īpaša struktūra mRNS 5’- galā • Nepieciešams translācijai eikariotos • Pievieno kepinga enzīms un metiltransfreāzes • Norisinās vienlaicīgi ar transkripciju • Apzīmējums: m7Gppp 5’- 5’ saite ar 3 fosfātu grupām Pirmo 2 nukleotīdu metilēšana
Poliadenilēšana • Gēnu 3’ galos ir t.s. poly-A signālsekvence: AAUAAA(N)50(G/U)bagāts rajons • Sasniedzot poly-A signālu : • 1. Transkripcija tiek terminēta • 2. RNS tiek sašķelta 10-35 nukleotīdus pēc AAUAAA sekvences • 3. Poly-A polimerāze pie RNS pievieno aptuveni 200 adenilātus • Poliadenilēšana ir nepieciešama: • 1) transkripcijas terminācijai • 2) mRNS transportam uz citoplazmu • 3) translācijai • 4) mRNS aizsardzībai pret eksonukleāzēm
Splaisings: intronu izšķelšanaKonsensus sekvence splaisa saita tuvumā YYYY
Splaisinga molekulārais mehānisms • Splaisings notiek ar divu transesterifikācijas reakciju palīdzību • Rezultātā izšķeļas «P» veida lariātaintrons • Reakciju katalizēsplaisosoma – liels proteīnu un RNS komplekss
mRNS nukleārais transports • Process, kurā mRNS tiek transportēta uz citoplazmu m7Gppp mRNS mRNS eksporteris A A A AA Nukleoplazma Kodola membrāna Nukleārā pora Citoplazma
Translācijas fāze • Gēna sekvence: • AGGTCATGTCTAGGTATGCCC • 1. fāze: • AGGTCATGTCTAGGTATGCCC • Arg Ser Cys Leu Gly Met Pro • 2. fāze: • A GGTCATGTCTAGGTATGC CC • Gly His Val ---- Val Cys Pro • 3. fāze: • AG GTCATGTCTAGGTATGCC C • Val Met Ser Arg Tyr Ala • Vēl 3 fāzes ir uz otrā dsDNA pavediena
Trīs galvenie RNS veidi • mRNS – translācijas matrica • rRNS – ribosomu sastāvdaļa • tRNS – nolasa mRNS kodu
tRNS tRNS satur atbilstošās aminoskābes antikodonu – t.i. kodonam komplimentāru sekvenci
Aminoacil-tRNS sintēze Augstas enerģijas estera saite Aminoskābe (Phe) Phe un tRNSPhe savienošana Phe aminoacil-tRNS sintetāze Phe tRNS (tRNSPhe) Aminoacil-tRNS
Ģenētiskā koda nolasīšanas mehānisms tRNSPhe saistās ar Phe kodonu UUU Kopējais rezultāts: Phe atpazīst savu kodonu Aminoacil-tRNS mRNS
Ribosoma • Proteīnu biosintēzes mašinērija • 65% RNS, 35% proteīns • 4 dažādas RNS, 83 proteīni (eikariotiem) • Katalītisko aktivitāti veic RNS daļa
Translācijas faktori • Proteīni, kuri nav ribosomu sastāvā, bet kuri ir nepieciešami dažādos translācijas etapos – iniciācijā, elongācijā un terminācijā • Iniciācijas faktori (eIF1, eIF2...) • Elongācijas faktori (eEF1, eEF2) • Atbrīvošanas (angl. «release») faktori (eRF1, eRF3), nepieciešami translācijas terminācijai
Eikariotiskās ribosomas subvienības eIF6 un eIF3 neļauj 60S un 40S subvienībām priekšlaicīgi apvienoties
Iniciācija Pirmās aminoacil tRNS piesaistīšana Preiniciācijas komplekss
Preiniciācijas komplekss mRNS piesasitīšana Iniciācijas komplekss
Iniciācijas komplekss mRNS skanēšana un starta kodona atpazīsana