Download
1 / 33
340 likes | 497 Views
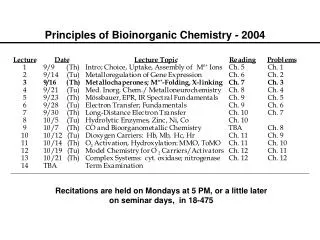
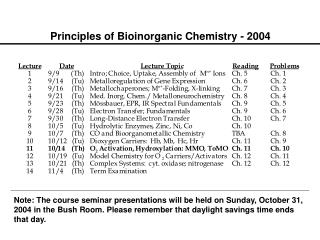
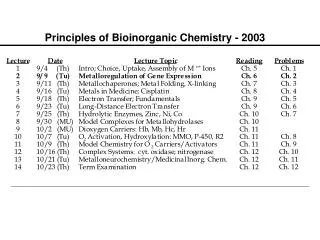
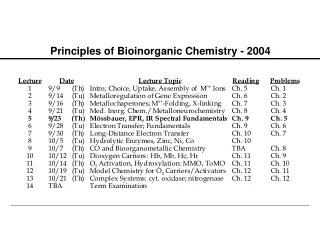
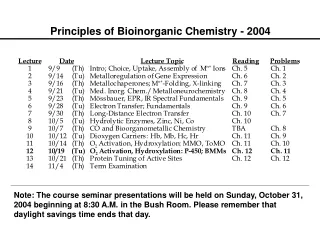
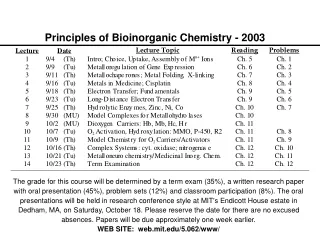
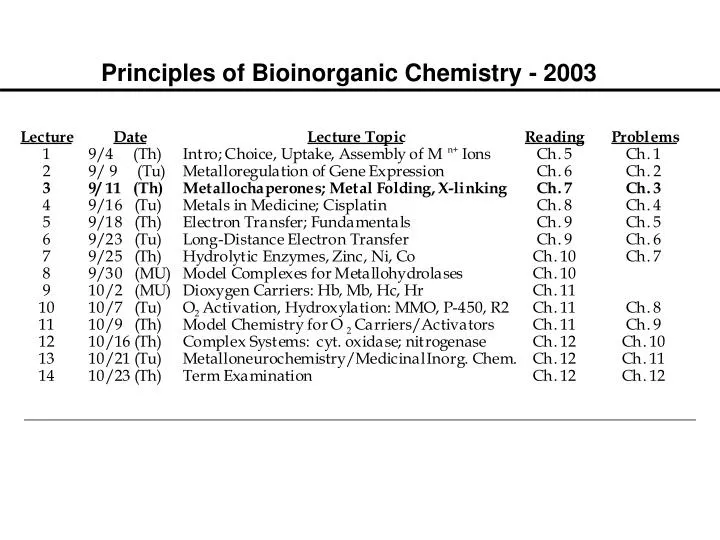
Principles of Bioinorganic Chemistry - 2003. Metalloregulation of Iron Uptake and Storage. Bacteria :

E N D
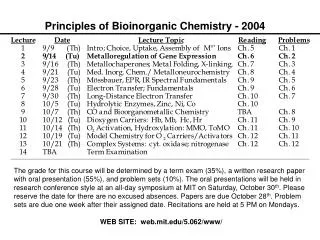
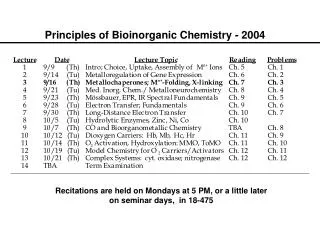
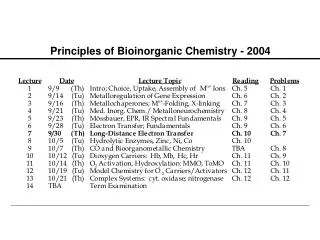
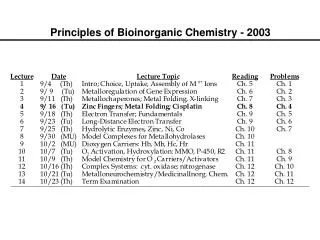
Metalloregulation of Iron Uptake and Storage Bacteria: A single protein, Fur (for iron uptake regulator), controls the transcription of genes involved in siderophore biosynthesis. Fur is a dimer with subunits of Mr 17 kDa. At high iron levels, the Fur protein has bound metal and interacts specifically with DNA repressing transcription. Mammals: Expression of ferritin and the transferrin receptor is regulated at the translational level.
Components of the Metalloregulatory System Fe IRP Iron-responsive protein (IRP) Stem-loop structure in the mRNA IRP
Regulation events High Fe, low TfR, high Ft Low Fe, high TfR, low Ft Fe IRP Message translated Message degraded Ferritin Transferrin IRP Message blocked Message translated
IRP1 is the Cytosolic Aconitase Contains an Fe4S4 Cluster Cluster assembled in protein, which then dissociates from mRNA Apoprotein stays associated with mRNA
Metallochaperones; Metal Folding PRINCIPLES: • Metallochaperones guide and protect metals to natural sites • Chaperone and target receptor protein structurally homologous • Metal-mediated protein structure changes affect transcription • Metal-mediated protein structure changes affect translation • Metal-induced protein structure changes also activate enzymes • Metal-induced bending of DNA affects function • Metal ionic radii and M–L water bridging are used to advantage ILLUSTRATIONS: • Copper insertion into metalloenzymes • Zinc finger proteins control transcription • Ca2+, a second messenger and sentinel at the synapse • Cisplatin, an anticancer drug
Copper Uptake and Transport in Cells The players: SOD, superoxide dismutase, a copper enzyme, a dimer containing two His-bridged Cu/Zn sites CCS, a copper chaperone for superoxide dismutase Lys7, the gene encoding yCCS in yeast; CCS and SOD1 co-localize in human tissue Ctr, family of membrane proteins that transport copper across the plasma membrane, delivering it to at least three chaperones: CCS, Cox17, Atx1 The puzzles: The total cellular [Cu] in yeast is 0.07 mM, none free How does copper find its way into metalloproteins? The implications: Mn, Fe, Zn have similar systems; understanding one in detail has implications for all
Two well characterized pathways Atx1 delivers Cu to transport ATPases in the secretory pathway, which translocates it into vesicles for insertion into multicopper oxidases such as ceruloplasmin Mutations in human forms of these ATPases lead to Menkes and Wilson diseases CCS delivers copper to Cu,Zn SOD Human Cu,Zn SOD is linked to ALS
Key Questions Address by Structural Bioinorganic Chemistry (Rosenzweig, O’Halloran, Culotta) What are the details of copper binding by ¨ these proteins, including stoichiometry and coordination geometry? How do these chaperones interact with their ¨ copper receptor proteins? What features of the copper binding and ¨ protein-protein interactions render each chaperone specific for its target protein?
Structure of the Hg(II) form of Atx1 Cys 15 Hg Hg(II) is exposed at the surface of the protein, which is reasonable for a protein that functions in metal delivery-- metal sites in enzymes are more buried. Hg(II) coordinated by the 2 cysteines. The apo protein has same structure but with a disulfide bonds between the cysteine residues. Cys 18 C N
More Details of the 1.2Å Structure, Active Site Cys 15 Thr 14 Ser 16 Val 12 2.33 Å Hg 2.34 Å Ser 19 Met 13 Cys 18 Lys 65 Ala 21
Structure of the Cu Hah1 Protein, the Human Homolog C N First copper chaperone structure with Cu bound The two molecules are primarily held together by the bound metal ion and some hydrogen bonding
Extended H-Bonding Interactions Stabilize the Structure T11B is conserved in most related domains. When it is not there it is replaced by His, which could serve the same function.
Postulated Mechanism for Metallochaperone Handoff of Copper to a Receptor Protein (O’Halloran, Rosenzweig, Culotta, 2000) HgAtx1 HgHah1 CuHah1 AgMenkes4
N 229CXC231 yCCS1 Crystal Structure • Domain I (Atx1-like) • metal binding • not essential • Domain II (SOD1-like) • target recognition C C20 C17 • Domain III • metal delivery • crucial Lamb, et al. Nature Struct. Biol. 1999, 6, 724-729
Dimer of Dimers Model + 54 kDa 32 kDa 86 kDa • SOD1 homodimer is very stable • yCCS and hCCS are dimeric in the crystal and in solution (yCCS under some conditions)
Heterodimer Model + 54 kDa 32 kDa 43 kDa • Structures indicate heterodimer formation is feasible • Heterodimer formation between different SOD1s • has been observed
Biophysical and biochemical studies of complex formation • According to gel filtration chromatography, dynamic light scattering, analytical ultracentrifugation, and chemical crosslinking experiments, yCCS and SOD1 form a specific protein-protein complex • The molecular weight of the complex, ~43 kDa, is most • consistent with a heterodimer • Higher order complexes, such as a dimer of dimers, were • not detected 86 kDa 43 kDa Lamb, et al. Biochem. 2000, 39, 14720-14727
Factors Affecting Heterodimer Formation • The heterodimeric complex formed with a mutant of SOD1 that cannot bind copper, H48F-SOD1, is more stable • Heterodimer formation is facilitated by zinc • Heterodimer formation is apparently independent of whether • copper is bound to yCCS • Heterodimer formation between Cu-yCCS and wtSOD1 in • the presence of zinc is accompanied by SOD1 activation • These data suggest that in vivo copper loading occurs via a • heterodimeric intermediate Lamb, et al. Biochem. 2000, 39, 14720-14727
Crystals of the yCCS/H48F-SOD1 heterodimeric complex P3221 a = b = 104.1 Å, c = 233.7 Å Solved by molecular replacement Lamb, et al. Nature Struct. Biol. 2001, in press.
Domain I SOD1 homodimer yCCS homodimer Domain III Domain II H48F-SOD1 monomer yCCS monomer
Domain I Domain II Domain III
Two heterodimers in the asymmetric unit Loop 7 Loop 7
Domain II Domain I C20 C17 C20 C229 C17 Domain III C231
C146 S-S subloop C57 C57 SO42-
C146 F48 C57 C229 C231
Mechanism of metal ion transfer • yCCS Domain I probably does not directly deliver the metal ion • yCCS Domain III is well positioned in the heterodimer to insert the metal ion • Transient intermonomer disulfide formation may play a role in yCCS function His 63 His 48 His 46 His 120 Cys 57 Cys 229 Cys 231
Metallochaperones; Metal Folding PRINCIPLES: • Metallochaperones guide and protect metals to natural sites • Chaperone and target receptor protein structurally homologous • Metal-mediated protein structure changes affect transcription • Metal-mediated protein structure changes affect translation • Metal-induced protein structure changes also activate enzymes • Metal-induced bending of DNA affects function • Metal ionic radii and M–L water bridging are used to advantage ILLUSTRATIONS: • Copper insertion into metalloenzymes • Zinc finger proteins control transcription • Ca2+, a second messenger and sentinel at the synapse • Cisplatin, an anticancer drug
Zinc Fingers - Discovery, Structures A. Klug, sequence gazing, proposed zinc fingers for TFIIIA, which controls the transcription of 5S ribosomal RNA. Zn2+ not removed by EDTA. 9 tandem repeats. 7-11 Zn/protein. Y or F – X – C – X2,4 – C – X3 – F – X5 – L – X2 – H – X3,4 – H – X2,6 C C C H H H H H The coordination of two S and 2 N atoms from Cys and His residues was supported by EXAFS; Zn–S, 2.3 Å; Zn–N, 2.0 Å. Td geometry. The protein folds only when zinc is bound; > 1% of all genes have zinc finger domains.
Structure of a Three Zinc-Finger Domain of Zif 268 Complexed to an Oligonucleotide Containing its Recognition Sequence
The Specificity of Zinc for Zinc-finger Domains Kd value: 2 pM 5nM 2mM 3mM Metal ion: Zn2+ Co2+ Ni2+ Fe3+