Download

1 / 39
420 likes | 896 Views
Effector Mechanisms of Cell-Mediated Immunity. Cell Mediated Immunity. Historically, immunologist have divided adaptive immunity into namely: CMI , which can be adoptively transferred only by viable T lymphocytes and

E N D
Cell Mediated Immunity • Historically, immunologist have divided adaptive immunity into namely: • CMI, which can be adoptively transferred only by viable T lymphocytes and • humoral immunity, which can be adoptively transferred with serum containing antibodies.
Cell Mediated Immunity • General responses by CMI, include: • Facilitate innate immune response to bacteria • Anti-viral • Anti-fungal • Anti-tumor • Transplantationrejection
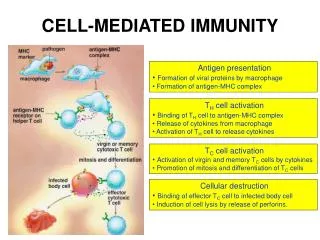
Cell Mediated Immunity • Many microbes have developed mechanisms that enable them to survive and even replicate within phagocytes, so the innate immunity is unable to eradicate infections by such microbes. • In CMI against phagocytosed microbes, the specificity of the response is due to T cells – but the actual effector function is mediated by the phagocytes.
CD4+ TH1 cells and CD8+ T cells recognize class II MHC-associated or class I MHC-associated peptide antigens of phagocytosed microbes, respectively, and produce cytokines (IFN-g, TNF) that activate the phagocytes to kill the microbes and stimulate inflammation.
Listeria monocytogenes • Protection to Listeria infection can be adoptively transferred by infusing T cells from an infected mouse into a naïve mouse. • This experiment was performed in 1950 by George Mackaness.
Delayed-type Hypersensitivity • There is a hypersensitivity condition that also demonstrates the T cell activation of macrophages. • Delayed-type hypersensitivity (DTH) is responsible for tissue injury due to Mf and pro-inflammatory cytokine release. • DTH will be discussed in the hypersensitivity lectures.
DTH Fig. 13-12
DTH Fig. 13-13
Cell Mediated Immunity • In a classical sense of CMI, CD8+ lymphocytes kill non-phagocytic cells infected with microbes.
Cell Mediated Immunity • CMI in response to helminthic parasites is mediated by TH2 cells that stimulate the production of IgE and activation of eosinophils.
Cell Mediated Immunity • Cell-mediated immune responses consist of the development of effector T cells from naïve cell in peripheral lymphoid organs, migration of these effector T cells and other leukocytes to sites of infection, through: • either cytokine-mediated activation of leukocytes to destroy microbes or • direct killing of infected cells.
Cell Mediated Immunity • Keep in mind that naïve T cells do not produce effector cytokines or the molecules to kill other cells. • The development of the effector T cells of CMI involves the sequence of antigen recognition, clonal expansion, and differentiation – as we have previously defined.
Cell Mediated Immunity • CD4+ cells may differentiate into subsets of effector cells that produce distinct sets of cytokines and therefore distinct effector functions. • These effector cell have been previously defined by us as TH1 and TH2.
TH1 and TH2 • The control of TH polarization is through the dendritic cell. • DC1 polarizes TH1 through IL-12 • DC2 polarizes TH2 through IL-4 • DC1 function appears to be through Toll-like receptors that bind bacterial DNA motifs (reviewed p 282-283). • DC2 function appears to be though increased levels of cAMP.
Toll-like receptors • Toll-like receptors (TLRs) are a family of pattern recognition receptors that are activated by specific components of microbes and certain host molecules. • INNATE RESPONOSE: They constitute the first line of defense against many pathogens and play a crucial role in the function of the innate immune system. • ADAPTIVE RESPONSE: TLRs were observed to influence the development of adaptive immune responses, through activating antigen-presenting cells, DC1.
Toll-like receptors (TLR) • TLRs are type I transmembrane proteins • TLR3 recognizes dsRNA, a viral product • TLR9 recognizes unmethylated CpG motifs frequently found in the genome of bacteria and viruses, but not vertebrates. • TLR7 recognizes small synthetic immune modifiers including imiquimod, R-848, loxoribine, and bropirimine, all of which are already applied or promising for clinical use against viral infections and cancers.
Toll-like receptors (TLR) • Plasmacytoid dendritic cells express TLR7 and TLR9, and respond to TLR7 and TLR9 ligands by producing a large amount of IFN-a. • TLR3, TLR7 and TLR9 play an important role in detecting and combating viral infections.
TH1 • TH1 • The polarization to TH1 can be stimulated by intracellular bacteria and viruses that infect macrophages. • A second pathway already described is microbe engagement of TLR. • A common feature of these infections is that they elicit innate immune reactions with the production of IL-12. • Enhancement of IL-12 production is through TH CD40L with APC CD40.
TH1 TH1 • IL-12 binds to CD4+ cells and activates STAT4, a transcriptional factor that promotes TH1 polarization. • IFN-g also induces T-bet, a transcriptional factor that enhances the TH1 polarization. • IFN-g stimulates further production of IL-12 by APCs and IL-12r on the lymphocyte.
TH1 and TH2 TH2 • The differentiation of antigen stimulated T cells to TH2 is dependent on IL-4. • IL-4 activates the transcriptional factor STAT6. • GATA-3 transcriptional factor, increases due to antigen presentation and enhances the transcription of TH2 cytokine gene transcription.
Effector Mechanisms: T-cells • Chapter 6 • Accessory Molecules of T-cells. • Adhesion molecules.
Integrin • A concept has evolved suggesting that T cells initiate antigen-independent adhesive interactions with apposing cells to scan the surface for specific antigen, followed by even stronger antigen-dependent adhesive interactions that would allow for specific activation of T-cell proliferation, cytokine production, or the delivery of a lethal hit to the target cell.
Integrins • Integrins are important for many different physiologic processes, including embryogenesis, thrombosis, wound healing, tumorigenesis and immune responses. • The integrin supergene family consists of a number of cell surface ab heterodimers. • The a and b chains are type I transmembrane glycoproteins with a single hydrophobic transmembrane domain, a short cytoplasmic tail, and an extracellular domain that associates noncovalently to form the heterodimer.
Integrins Page 120
Integrins • In the immune system, the most important integrins, those of the b1, b2, and b7 subfamilies, participate in T-cell migration and provide stimulatory signals for T-cell proliferation and effector functions.
Integrins • T-cell migration into tissues requires T-cell binding to and extravasation through endothelium, an integrin-dependent process. • Current models propose that chemokines deliver the critical biochemical signals that promote endothelial binding through the upregulation of integrin avidity on the T cell.
Integrins • Integrin-mediated events are also critical for T cells that participate in immune surveillance. • As resting T cells circulate through the blood, they adhere specifically to the specialized endothelium of the postcapillary venules of secondary lymphoid organs or HEVs and extravasate from the bloodstream into the underlying secondary lymphoid tissues.
Integrins • How do the adhesion molecules regulate their binding activity to allow both the attachment as well as detachment characteristics required for transmigration into and out of the inflamed endothelium and draining lymph nodes? • Lymphocytes have solved this problem by tightly regulating the affinity and avidity of integrin receptors.
Integrins • These conformational changes are dependent on the presence of specific divalent cations, which are bound by the extracellular domains of integrins. • Replacement of bound Ca2+ with Mg2+ for b2 integrins or Mn2+ for b1 integrins results in increased receptor affinity for their ligands