Download

1 / 53
620 likes | 1.22k Views
Ligand-Gated Ion Channels. Molecular Biophysics 28 September 2007. LGIC mediate fast synaptic transmission. LGICs are responsible for changing a chemical signal in the synapse (neurotransmitter) to either an inhibitory or excitatory post synaptic potential in the post synaptic cell.

E N D
Ligand-Gated Ion Channels Molecular Biophysics 28 September 2007
LGICs are responsible for changing a chemical signal in the synapse (neurotransmitter) to either an inhibitory or excitatory post synaptic potential in the post synaptic cell.
The EPSPs and IPSPs are summed from all of the dendrites, changing the membrane potential at the axon hillock. If the depolarization is high enough, an AP will be initiated.
Fast exchange of bath is needed to study ligand-gated ion channels FSU Neuroscience Website Paul Trombley
Families of Ligand-Gated Ion Channels • Cys-loop receptors • Nicotinic Acetylcholine receptor • GABAA and GABAC Receptors • Glycine Receptor • 5-HT3 Receptor • Ionotrophic Glutamate Receptors • NMDA • AMPA • Kainate • P2X Receptors Kandel, Schwartz & Jessel, Principles of Neural Science 4th Ed. (2000)
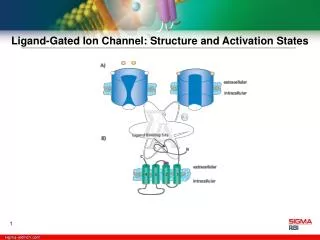
Cys Cys Cystine-Loop Superfamily of Ligand-Gated Ion Channels • Heteromeric or homomeric pentamers • Characterized by a large N-terminal loop cross-linked by cystine bridges • Each subunit is made up of 4 membrane spanning helices • The large intracellular M3-M4 linker is the site for many cytoskeletal protein-protein interactions. • M2 lines the pore Keramidas et al., 2004. Ashcroft 2000
nAChR • Activated by Acetylcholine and Nicotine • µs activation times • Blocked by curare and some general anesthetics. • Non-selective cation channel including sodium, potassium and calcium. • Isolated from Torpedo marmorata and visualized by N. Unwin and colleagues in the mid 1980’s
It’s pentameric structure consists of 2 a subunits and a mixture of b, d, and g subunits. Ligand binding domain Pore lined by TM2 Intracellular M3-M4 linker Unwin’s 2005 4 resolution electron microscopy structure.
Open and closed state of the channel at the gate is different by 3 Van der Waal’s surface representation at the gate. Unwin 2003
nAChR kinetics are dependent on subunit composition. Desensitization Giniatullin et al 2005
Cystine-Loop Superfamily of Ligand-Gated Ion Channels 5-HT3 Receptor
5-HT3 is a non-selective cation channel and is sensitive to curarie Yan et al. 1999
Homomultimers and heteromultimers of 5-HT3A and 5-HT3B Receptor Subunits produce channels with different characteristics. Peters et al 2005
Arginine residues with in the cytoplasmic domain strongly influence conductance of the 5-HT3 receptor Electrostatic potential surface representation Peters et al. 2004 Peters et al. 2005
Cystine-Loop Superfamily of Ligand-Gated Ion Channels Glycine and GABA Receptors
Gate Region Basic Residues Ion Selection: Chloride Channels Modified from Keramidas et al., Prog. Biophys. Mol. Biol.86: 161 (2004) Slide from Blitzer, Teaching Resource, Science’s STKE 2005
GABA Receptor SubunitComposition Two GABA Binding Sites ata-bInterfaces Benzodiazepine Site at a-g Interface GABA = Gamma-aminobutyric Acid Katzung (Ed.) Basic & Clinical Pharmacology, Lange (2004) Slide from Blitzer, Teaching Resource, Science’s STKE 2005
Probability of Opening Open Time Benzodiazepines and Barbiturates EnhanceGABAA Currents Through Different Mechanisms Twyman et al (1989) Ann. Neurol.25: 213-220 (1989) Slide from Blitzer, Teaching Resource, Science’s STKE 2005
GlyR Betz and Laube 2006
Cys Cys
Characterization of the Chimeric channel Expressed in Hek 293 cells Whole-cell patch recording of macroscopic chloride currents Functional Ca2+ potentiation site of the ECD ACh gates the channel Reversal potential shifted closer to Na when Cl cdriving force is removed Grutter et al 2005
Activation is slowed in the 7/Cly chimeria possibly due to mismatched interactions of the poorly conserved Cys-loop of nAChR and the M2-M3 linker of the Glycine receptor.
Replacement of the nAChR Cys-loop with the Glycine R Cys-loop speeds activation Grutter et al 2005
Cys-loop / M2-M3 linler interactions are important for activation Kinetics and they are receptor specific. Activation of Glycine R w/cys-loop point mutations to nAChR specific residues Slow activation of Chimeric Chimeric channel w/2-3 linker of nAChR WT Glycine R activation Grutter et al 2005
How many subunits make up an ionotrophic glutamate receptor?
Determination of binding sites by single channel electrophysiology • AMPA receptor composed of GluR6/GluR3 chimeric channel expressed in HEK293 cells • Form homomultimers • No desensitization • The assumptions • # of binding sites = # of subunits • Binding sites must be equivalent
These channels have 3 conductances and a closed state. Quisqualate = AMPA Receptor agonist NBQX = high affinity AMPA Receptor antagonist MNQX = lower Affinity AMPA Receptor antagonist Cyclothiazide = blocks inactivation Rosenmund et al 1998
The relative frequency current amplitude histogram should shift in a predictable manner with increasing concentration of agonist if the states observed are due to different #’s of bound ligand.
Dwell time analysis for each transition state indicates 4 subunits 2 components
Ionotrophic Glutamate Receptors NMDA Receptors
NMDA Receptors • NMDA = N-methyl-D-aspartic acid • Made up of at least 1 NR1subunit and a combination of NR2A-D and NR3A-B • Permeable to K+, Na+, Ca2+ • High conductance • Activate slowly • Desensitize slowly & incompletely • Prolonged Ca2+ influx in the face of sustained glutamate release
Different combinations of NMDA subunits produce channels with an array of kinetics Cull-Candy et al 2001
NMDA receptor and Mg2+ • Blocks channel at rest • Depolarization --> Mg2+ ion leaves the pore • Glu + depolarization = Coincidence Detector • Other channel blockers: PCP, ketamine, MK801 Zigmond et al. 1999
Ionotrophic Glutamate ReceptorsAMPA and Kainate Receptors • Activate rapidly • Desensitize within a few milliseconds • Kainate – GluR5-7, KA1-2 • AMPA – GluR1-4 • With GluR2 subunit: permeable only to K+ and Na+ • Without GluR2 subunit: Ca2+-permeable • AMPA = alpha-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid
Isolating AMPA-R and NMDA-R Currents With Selective Blockers Nestler, Hyman, & Malenka, Molecular Neuropharmacology McGraw-Hill (2001) Slide from Blitzer, Teaching Resource, Science’s STKE 2005
P2X Receptors • Gated by extracellular ATP • Trimeric arrangement determined by crosslinking and agonist binding studies • 7 subtypes, heteromultimers produce a variety of kinetic outcomes • Do not contain common ATP consensus motifs (ie. Walker motif) • M1 is involved in gating, M2 lines the pore • Intracellular N and C termini are important for protein-protein interactions
P2X receptors are permeable to both Na+ and Ca2+ and have a wide variety of kinetics P2X6 is silent but can be expressed with other subunits to modulate their kinetics Egan et al. 2006
hP2X1 • Cystine point mutations of S286-I329 • Oocyte expression, two-electrode voltage-clamp • Hek293 expression, whole-cell patch-clamp
Potency shift due to agonist binding and/or channel gating changes. oocytes Roberts and Evans 2007
Some mutations that do affect ATP potency, have decreased binding efficiency but not all Protein expression is not different across mutants ATP binding is decreased in 4 of the mutants in 32P 2-azido ATP /UV cross linking studies Roberts and Evans 2007
Decreased EC50 Increased EC50 Some mutations slow activation and desensitization in conjunction with or regardless of ATP potency changes. Roberts and Evans 2007
Addition of a charge to some mutated residues modulate peak current magnitude. MTS compounds forms disulfide bonds with the side chain of cystine when exposed MTSEA adds a positive charge WT P2X1 does not have an exposed cystine in the region in question MTSES adds a negative charge