Download

1 / 36
360 likes | 511 Views
H2A.Z-Mediated Localization of Genes at the Nuclear Periphery Confers Epigenetic Memory of Previous Transcriptional State. Donna Garvey Brickner, Ivelisse Cajigas, Yvonne Fondufe-Mittendorf, Sara Ahmed, Pei-Chih Lee, Jonathan Widom, Jason H. Brickner (PI) Northwestern University.

E N D
H2A.Z-Mediated Localization of Genes at the Nuclear Periphery Confers Epigenetic Memory of Previous Transcriptional State Donna Garvey Brickner, Ivelisse Cajigas, Yvonne Fondufe-Mittendorf, Sara Ahmed, Pei-Chih Lee, Jonathan Widom, Jason H. Brickner (PI) Northwestern University
Background – localization of chromatin at the nuclear periphery • In metazoan cells, heterochromatin and developmentally repressed genes localize at the periphery • In yeast, peripheral localization of telomeres and the mating type loci promotes silencing • In yeast and mice, some genes are recruited to the periphery when transcriptionally activated • It is thought that peripheral localization may be a mechanism of efficiently coupling transcription with export • Periphery tethering points can serve as hard boundaries to limit the spreading of heterochromatin formation
Background – INO1 and GAL1 • GAL1 - Galactokinase, phosphorylates alpha-D-galactose to alpha-D-galactose-1-phosphate in the first step of galactose catabolism • activated by growth in galactose rather than glucose • INO1 - encodes inositol 1-phosphate synthase, the enzyme that catalyzes the conversion of glucose 6-phosphate to inositol 1-phosphate • activated by inositol starvation • Because of their easy transcriptional activation or repression through choice of growth medium, they are used here as convenient reporter genes
Technique 1 – quantitative assay for subnuclear localization of genes – Lac tagging • A series of lac repressor-binding sites (lac operators) are introduced adjacent to a gene of interest by homologous recombination… Gene ORF L-O L-O L-O L-O • …In yeast strains expressing a GFP-tagged lac repressor, introduced on a plasmid LacI– GFP LacI– GFP LacI– GFP LacI– GFP Gene ORF L-O L-O L-O L-O
Technique 1 – quantitative assay for subnuclear localization of genes – Lac tagging • For visual reference, the nuclear envelope protein Sec63 is tagged with myc by similar molecular genetic methods • Cells are fixed, fluorescently labelled myc antibodies are introduced, and both these antibodies and GFP are visualized by fluorescence microscopy Nuclear envelope Sec63 Sec63 myc myc Sec63 myc LacI– GFP LacI– GFP LacI– GFP LacI– GFP Gene ORF L-O L-O L-O L-O Sec63 myc Sec63 myc
Question 1 – does transcriptional activation induce periphery recruitment? • Lac tagging method used to count the proportion of cells in which GAL1 or INO1 localize to the periphery before and after transcriptional activation is induced by glucose or inositol starvation • Dynamic range of the experiment: URA3, which is randomly distributed in the nucleus, co-localizes with Sec63-myc in 25% of cells – this is the minimum/baseline. INO1, artificially tethered to the nuclear envelope, co-localizes in 80% of cells – this is the maximum/ceiling Cell Population INO1 – uninduced - - - INO1 – induced - - - - - GAL1 – uninduced - - - GAL1 – induced - - - - - Percent of cells co-localizing with Sec63-myc 31 +/- 1 % 60 +/- 5 % 35 +/- 1 % 70 +/- 2.5 %
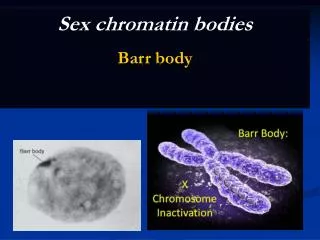
From: H2A.Z-Mediated Localization of Genes at the Nuclear Periphery Confers Epigenetic Memory of Previous Transcriptional State Brickner DG, Cajigas I, Fondufe-Mittendorf Y, Ahmed S, Lee PC, et al. PLoS Biology Vol. 5, No. 4, e81 doi:10.1371/journal.pbio.0050081 The proportion of cells demonstrating INO1 periphery localization before and after induction by inositol starvation Sec63-myc, which shows the nuclear envelope and ER, is in red. LacI-GFP, here associating with Lac operator repeats joined to the INO1 gene, is in green. This demonstrates a cell that would not be counted as demonstrating INO1 periphery localization, and a cell that would.
Technique 2 – Real-Time PCR A cell strain of interest is grown in the appropriate conditions, then the cells are harvested and lysed to expose their mRNA The mRNA is converted to cDNA by incubation with reverse transcriptase A set of primers appropriate to the gene of interest is designed, and the PCR reaction is carried out in a special machine in the presence of a fluorescent probe standard PCR schematic http://www.ncbi.nlm.nih.gov/books/bv.fcgi?highlight=PCR&rid=mboc4.figgrp.1590
The probe contains a fluorescent dye along with a quencher The advancing polymerase, encountering the probe, cleaves it, separating the dye from the quencher and increasing the fluorescent signal The increase in fluorescent signal intensity is therefore proportional to the number of probes bound, i.e. the number of copies present If more cDNA copies are initially present, the signal increases at a faster rate, and initial mRNA concentration is thus indirectly measured Technique 2 – Real-Time PCR http://www.servicexs.com/plaatjes/TaqMan_RT-PCR_assay.jpg
Question 2 – How fast does periphery localization occur relative to transcriptional activation – could one process be driving the other? • INO1 and GAL1mRNA expression, following induced transcriptional activation, was quantified at various time points using real time PCR • ACT1, a constitutively expressed gene with fairly constant cellular mRNA levels, was used as a control – absolute mRNA levels were not quantified, but rather relative levels as compared with ACT1 • Peripheral localization (measured as a percentage of cells) was assessed at various time points by the lac tagging method • These two parameters were compared linearly, and with the mRNA expression levels plotted on a log scale
Question 2 – How fast does periphery localization occur relative to transcriptional activation B: INO1 C: GAL1 Left side: Both parameters plotted linearly Right side: peripheral localization plotted linearly, mRNA expression plotted on log scale • A rapid increase in INO1 and GAL1 mRNA expression was observed following induction by nutrient starvation • This increase correlated with a rapid recruitment of both gene loci to the nuclear periphery • The maximum rate of mRNA accumulation for both INO1 and GAL1 occurred after nuclear periphery localization was largely complete
Question 3 – Do the genes remain localized to the periphery after transcription becomes repressed? • Repressed transcription by adding inositol to INO1-expressing cells and glucose to GAL1-expressing cells • Monitored mRNA expression and peripheral localization by the same procedures as above • mRNA expression for both strains returned to the repressed level within 30 minutes • Peripheral localization, however, persisted for more than two hours in both cases
Question 3 – Do the genes remain localized to the periphery after transcription becomes repressed? The INO1 and GAL1 gene loci remain localized at the nuclear periphery more than two hours following transcriptional repression, despite a rapid return to repressed-level mRNA levels • Cells are capable of rapidly changing the distribution of these loci, as evidenced by their rapid peripheral localization following gene activation, as well as the high diffusion constant for the repressed GAL1 locus (reported by Cabal et. al. 2006) • Therefore, an active mechanism must be employed to retain these genes at the periphery
Question 4 – Is continued INO1 periphery localization merely due to low but sufficient levels of transcription? • INO1 may remain at the periphery because adding inositol to the cells is not sufficient to achieve a full repression effect, and INO1 is still being transcribed • To test this, mutant strain rbp1-1 was used. These cells lose RNA polymerase II function above the restrictive temperature. • Cells starved with inositol to activate INO1 transcription and periphery localization cells raised to the restrictive temperature, inactivating RNA polymerase II and completely ending INO1 transcription INO1 localization was monitored
Question 4 – Is continued INO1 periphery localization merely due to low but sufficient levels of transcription? Right panel: INO1 mRNA levels predictably fall after raising the cells to the restrictive temperature and inactivating RNA polymerase II Left panel: INO1 peripheral localization at restrictive temperature Black dots: wild type strain Black squares: temperature sensitive RNA polymerase mutant White circles: mutant grown in repressed conditions Conclusion: Once activated, continued transcription is not required to maintain INO1 at the nuclear periphery
Question 5 – Is actual transcription required to establish INO1 periphery recruitment? • Next, it was tested whether active transcription by RNA polymerase II, in addition to inositol starvation, is required for INO1 to attain periphery localization • To test this, the rbp1-1 mutant was • Raised to the restrictive temperature, inactivating RNA polymerase II • Then starved of inositol • Despite being raised to the restrictive temperature prior to induction, the rpb1-1 mutant exhibited INO1gene localization at the nuclear periphery to the same extent as in the wild type strain. • These results indicate that the mechanism responsible for sensing inositol starvation and recruiting INO1 to the nuclear periphery is independent of the activity of RNA polymerase II and the transcriptional machinery
RECAP • INO1 and GAL1, two commonly used reporter genes, are recruited to the nuclear periphery following the introduction of the cells to activating conditions • Periphery localization and increase in mRNA expression are concurrent, with the highest level of increase in mRNA expression occurring after periphery localization is complete • These genes remain at the periphery long after the cells are returned to repressing conditions and mRNA expression returns to repressed levels • Transcription by RNA polymerase II is not required either to recruit INO1 to the nuclear periphery or to retain it there
Question 6 – Is INO1 and GAL1 periphery localization inherited? • To determine whether gene localization at the periphery is a heritable (and thus, likely, epigenetic based) state, cells were • Activated • Repressed • Maintained in logarithmic growth by dilution with growth medium • After transcription is repressed, INO1 and GAL1 are maintained at the nuclear periphery, and this state is inherited for several generations • 3-4 generations for INO1 • 65 generations for GAL1 (!) • This suggests that localization is maintained through epigenetic mechanisms
Question 7 – Does the peripheral localization of short-term repressed INO1 and GAL1 facilitate more rapid reactivation? Short-term repressed cell populations experience much more rapid reactivation of GAL1 mRNA expression than long-term repressed populations Short term repressed: Galactose glucose galactose Long term repressed: Glucose galactose Growths carried out for several cell generations in each medium
Question 7 – Does the peripheral localization of short-term repressed INO1 and GAL1 facilitate more rapid reactivation? Short-term repressed cells experience slower reactivation of INO1 mRNA expression following the shift to activating conditions, due to the specifics of activation and the molecules involved … …However, mutantslacking a nuclear pore complex component necessary for peripheral tethering experience even slower reactivation, suggesting that peripheral localization again has a positive effect on reactivation rapidity
Question 8 – Can nuclear periphery localization alone promote rapid transcriptional activation regardless of transcriptional history? To answer this question, a cell line was created in which the INO1-lac operator array was supplemented with a nuclear envelope-targetting motif called FFAT • Tethering was found to have no effect on steady-state levels of INO1 mRNA expression in any growth medium conditions • However, artificially tethered INO1 experienced a more rapid increase in mRNA expression upon inositol starvation than untethered (wild-type) INO1
RECAP • INO1 and GAL1, two commonly used reporter genes, are recruited to the nuclear periphery following the introduction of the cells to activating conditions • Periphery localization and increase in mRNA expression are concurrent, with the highest level of increase in mRNA expression occurring after periphery localization is complete • These genes remain at the periphery long after the cells are returned to repressing conditions and mRNA expression returns to repressed levels, and this localization is inherited for several generations • Transcription by RNA polymerase II is not required either to recruit INO1 to the nuclear periphery or to retain it there • The peripheral localization of short-term repressed genes facilitates more rapid transcriptional activation • This localization alone is sufficient to promote rapid activation, regardless of transcriptional history
Technique 3 – Nuclease digestion of chromatin to reveal nucleosomal positioning • Cells are permeablized and treated with nuclease to digest unprotected DNA • RT Q-PCR and many primer sets are used to determine relative concentrations of overlapping DNA sequences • High concentrations indicate extensive nucleosomal protection • Well-known nucleosomal and non-nucleosomal sites are used as controls
Question 9 – Is there a difference between short-term and long-term repressed INO1 genes in terms of nucleosomal positioning or nucleosomal protection? • Two stably positioned nucleosomes identified in INO1 – one at the promoter, and one between the promoter and the ORF • There was no change in the positioning of these nucleosomes between short-term and long-term repressed INO1 • There was, however, a decrease in nucleosomal protection in short-term repressedINO1 • Decreased protection can indicate either an increased in the proportion of cells lacking nucleosomes, or a decrease in nucleosomal stability…and because of something about the positioning of the two nuclesomes…they must contain…
H2A.Z! Jason Brickner, as portrayed by Edward Norton
H2A.Z • A nucleosome containing a variant histone component encoded by the htz1 gene • Has been shown to form boundaries preventing the spread of heterochromatin • Inclusion at gene promoters negatively correlates with transcriptional frequency • Htz1 is deposited at promoters during repression, but is not required to establish the repressed state • It is thought that Htz1 inclusion facilitates rapid activation because Htz1 containing nucleosomes are less stable and more easily ejected from promoter regions than their H2A-containing counterparts • In an in vitro chromatin system, far lower ionic strength was required to separate Htz1 from DNA than to separate H2A or H3
Question 10 - is INO1 or GAL1 transcriptional activation altered in htzΔ1 mutants? Loss of H2A.Z led to a significant delay in short-term repressed Ino1 and Gal1 reactivation Loss of H2A.Z had no effect on long-term repressed Ino1 and Gal1 reactivation
Question 11 – Are there more H2A.Z nucleosomes associated with short-term repressed INO1 promoters than with long-term repressed promoters? Chromatin immunoprecipitation, using an antibody that recognizes htz1, was used to precipitate H2A.Z and analyze the DNA sequences to which it is bound In long-term repressed or transcriptionally active cells, the bound DNA was not enriched for the INO1 promoter In short-term repressed cells, there was a strong enrichment for the INO1 promoter This suggests that H2A.Z is incorporated into the INO1 promoter region following repression
Question 12 – Are htz1Δ cells deficient in peripheral localization of the Ino1 gene? htz1Δ cells are not deficient in localizing Ino1 to the periphery when actively transcribing… …but they are deficient in maintaining Ino1 at the periphery following repression
RECAP • H2A.Z nucleosomes are incorporated into the INO1 promoter following repression • htz1Δ mutant cells exhibit slower reactivation of short-term repressed Ino1 and Gal1 but are unaffected as far as reactivation of these genes after long-term repression • htz1Δ mutant cells are able to recruit Ino1 to the nuclear periphery following activation, but are unable to retain the gene at the periphery following repression Gene repression Peripheral localization H2A.Z incorporation Rapid reactivation following short-term repression The correlations are striking, but causal relationships are unclear
MODEL • Short-term and long-term repressed INO1 and GAL1 are distinguished by their localization, transcriptional history, and kinetics of activation • Peripheral localization of INO1 is necessary and sufficient to promote more rapid activation, regardless of transcriptional history • H2A.Z incorporation is necessary to promote more rapid activation • Unanswered question: • Is the easy dissociation of H2A.Z nucleosomes the mechanism, or is H2A.Z important only in promoting peripheral tethering, which itself provides the optimal environment for reactivation? • I would have liked to see an experiment in which genes in htz1Δ mutants were artificially tethered to the periphery
MODEL A MODEL B Recruitment to the periphery promotes H2A.Z incorporation, which allows for rapid reactivation due to the ability of H2A.Z to easily dissociate from DNA Incorporation of H2A.Z nucleosomes promotes peripheral localization, and the periphery provides an optimal environment for rapid gene activation
Significance of this work • Demonstrates a case in which cells inherit not a transcriptional state, the usual epigenetic paradigm, but rather transcriptional information • Demonstrates a novel role of H2A.Z, that of ‘marking’ recently repressed genes and promoting their peripheral retention and rapid reactivation
Hypothesis on why short-term repressed INO1 takes longer to reactivate, in case anyone asks • Phosphatidic acid is combined with inositol to produce phosphatidylinositol, and is the activating molecule for INO1 • Low inositol concentration leads to a buildup of phosphatidic acid, which is responsible for the activation of INO1 • When cells are starved of inositol to cause expression, Ino1p is produced which in turn produces inositol • When cells are then grown in inositol to initiate repression, a significant amount of Ino1p remains in the cell, which continues to produce inositol and perpetuates the activity of the pathway that depletes phosphatidic acid • This produces a lag time before enough phosphatidic acid can accumulate to activate Ino1 expression again