Download
1 / 47
490 likes | 700 Views
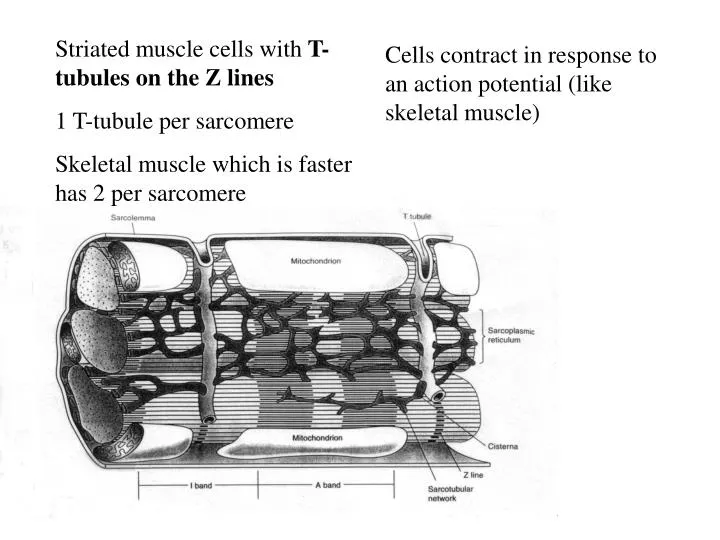
Striated muscle cells with T- tubules on the Z lines 1 T-tubule per sarcomere Skeletal muscle which is faster has 2 per sarcomere. Cells contract in response to an action potential (like skeletal muscle) . Many mitochondria. Site of aerobic metabolism primarily of FFA and Lactate.

E N D
Striated muscle cells with T- tubules on the Z lines 1 T-tubule per sarcomere Skeletal muscle which is faster has 2 per sarcomere Cells contract in response to an action potential (like skeletal muscle)
Many mitochondria. Site of aerobic metabolism primarily of FFA and Lactate Only ~15% of metabolism is glucose
Desmosomes transmit mechanical force from cell-to-cell. They are sort of “molecular rivets” Action potentials propagate through gap junctions. Striated appearance comes from overlapping thick (myosin) and thin (actin) filaments
Titin Muscle
Myosin II molecules are double trimers of a heavy myosin chain, a regulatory light chain and an alkali light chain
Contraction results from X-bridge interaction between actin and myosin. Energy is supplied by ATP. ATP is required to break X-bridges ATP hydrolysis occurs when the head is cocked
Tropomyosin inhibits X-bridge formation between actin and myosin. Binding of Ca++ to troponin C interrupts that inhibition and allows X-bridge cycling.
Phase 2 is the plateau Due to Ca++ influx through slow (L-type) calcium channels. They are called slow because they stay open for about 300 ms Prolongs the action potential to 300 ms to maintain force generation and create a long refractory period to insure a diastolic relaxation period between beats.
The action potential is carried to the interior of the cell by the transverse tubules. That allows rapid activation of the interior of these big (25u) cells.
Ca++ coming out of the transverse tubules (trigger calcium) binds to ryanodine receptors and cause Ca++ stored in the sarcoplasmic reticulum to be released into the cytosol. 1 Skeletal muscle only Cardiac muscle only 2 3
Some calcium enters the cell through the slow Ca++ channels during phase 2. This is called Trigger Ca++. Extracellular Fluid ++ Ca + K Sarcolemma ATP Pump Ex L type channel Cytosol + ++ Na + Ca Na ++ Ca Trigger ATP SERCA ++ Ca Sarcoplasmic reticulum Ryanodine receptor Sliding filaments
Trigger Ca++ binds to the ryanodine receptors on the sarcoplasmic reticulum (SR). Extracellular Fluid ++ Ca + K Sarcolemma ATP Pump Ex L type channel Cytosol + ++ Na + Ca Na ++ Ca Trigger ATP SERCA ++ Ca Sarcoplasmic reticulum Ryanodine receptor Sliding filaments
Extracellular Fluid ++ Ca + K Sarcolemma ATP Pump Ex L type channel Cytosol + ++ Na + Ca Na ++ Ca Trigger ATP SERCA ++ Ca Sarcoplasmic reticulum Ryanodine receptor Sliding filaments Most of the calcium that activates the muscle comes from the SR. This is termed calcium-induced calcium release. The Ryanodine receptors are actually Ca++ channels. Calcium binding causes them to open and release Ca++ that is stored in the SR
Heart is very sensitive to serum Ca++. Low serum calcium results in loss of trigger calcium and a weak heart beat
Extracellular Fluid ++ Ca + K Sarcolemma ATP Pump Ex L type channel Cytosol + ++ Na + Ca Na ++ Ca Trigger ATP SERCA ++ Ca Sarcoplasmic reticulum Ryanodine receptor Sliding filaments After the action potential subsides calcium is pumped back into the SR by a calcium ATPase,called SERCA.
Much of the calcium stored in the SR is bound to calsequestrin and calreticulum thus increasing the SR’s capacity. Ca++ has low solubility and would precipitate in the SR without these proteins.
Calcium is moved out of the cell by the Na/Ca exchanger. To maintain a steady state as much Ca++ leaves the cell over one cycle as entered. The exchanger gets its energy from the sodium gradient. A small amount of calcium is also pumped out by a sarcolemmal calcium ATPase (pump) Extracellular Fluid ++ Ca + K Sarcolemma ATP ATP Pump Pump Ex L type channel Cytosol + ++ Na + Ca Ca++ Na ++ Ca Trigger ATP SERCA ++ Ca Sarcoplasmic reticulum Ryanodine receptor Sliding filaments
About 75% of the cytosolic Ca++ comes from the SR with each beat
Insufficient Ca++ is released with each action potential to cover all troponin binding sites. Therefore, increasing the cytosolic calcium concentration increases the force of contraction The strength of the beat is proportional to the amount of calcium stored in the SR. SERCA competes with surface exchangers for Calcium during phase 4 and anything that promotes more calcium into the SR increases the force of contraction.
Extracellular Fluid ++ Ca + K Sarcolemma ATP Pump Ex L type channel Cytosol + ++ Na + Ca Na ++ Ca Trigger ATP SERCA ++ Ca Sarcoplasmic reticulum Ryanodine receptor Sliding filaments Decreasing the sodium gradient by the sodium pump inhibitor digitalis causes less calcium to leave via the exchanger and consequently more to enter the SR. That increases force of contraction.
Extracellular Fluid ++ Ca + K Sarcolemma ATP Pump Ex L type channel Cytosol + ++ Na + Ca Na ++ Ca Trigger ATP SERCA ++ Ca Sarcoplasmic reticulum Ryanodine receptor Sliding filaments Because digitalis also poisons the Na+/K+ ATPase in muscle and nerves it can easily lead to arrhythmias, paralysis and death. low therapeutic index
Increasing the heart rate increases the time spent in phase 2 (Ca++ entry) and decreases time spent in phase 4 (Ca++ exit). This causes more Ca++ to accumulate in the SR. Hence increasing the heart rate increases force of contraction
How can we vary the force of contraction in cardiac muscle? • Increase muscle length • Increase the duration of the active state (skeletal only) • Increase contractility (cardiac only)
Change the resting muscle length (preload). As in skeletal muscle the force of contraction is very dependent on the geometry of the overlap between the thick and thin filaments Starling’s law of the heart: The more the heart is filled during diastole the more forcefully it will contract during systole
The top figure is actually for skeletal muscle because the relaxed tension is still zero at the peak of the contracted curve (LMAX). Contrast that to the curve in in the lower panel. The high resting tension is needed to resist over-extension of the muscle by the diastolic pressure.
Active Most of the length-dependency comes from the mechanical geometry between the thick and thin filaments.
Change the contractility Inotropy A change in contractility is defined as a change in force of contraction not related to a change in length. A change in contractility can be seen as a shift in the “contracted” curve either up (an increased contractility) or down (a decreased contractility)
Sympatheic nerve activity is the usual cause of an increased contractility. Heart failure occurs when contractility is pathologically reduced Contractility is hard to measure in patients
The force of contraction is influenced by catecholamines through the beta 1 receptors
Beta 1 receptors are coupled to stimulatory G protein. They act to increase cAMP in the muscle cells.
cAMP activates a kinase, Protein Kinase A (PKA) PKA puts a phosphate group on specific target proteins.
Protein kinase A (PKA) phosphorylates L-type Ca++ channels (opens them) allowing more Ca++ to enter the cell during phase 2.
In 1972 Michihiko Tada found that the ATPase activity of a preparation of cardiac muscle sarcoplasmic reticulum could be increased by adding cAMP. They later found that was dependent on a protein they named phospholamban Michi Tada 1938-2005
Tada found that phospholamban normally inhibits SERCA. Phosphorylation by PKA inhibits the inhibition (Activates SERCA) Ser16 cAMP PKA Phospholamban SERCA Ca++
Protein kinase A (PKA) Phosphorylates phospholamban which in turn increases SERCA activity causing more Ca++ to be sequestered in the SR
Cytosolic Ca++ binds calmodulin which then activates CAM kinase. CaM-kinase phosphorylates phospholamban at Thr17 and also increases SERCA activity. Ser16 cAMP PKA Thr17 SERCA Phospholamban Calmodulin CaM-kinase + Ca++ CaM Ca++
CaM-kinase acts with sympathetic stimulation through elevated Ca++ to amplify the response.
Activation of SERCA by phosphorylating phospholamban increases force of contraction by putting more releasable Ca++ in the SR. Phospholamban phosphorylation also speeds the rate of relaxation by removing Ca++ from the cytosol more quickly (+Lusitropic effect). That promotes diastolic filling. Phospholamban actually inhibits SERCA Mice in which phospholamban has been ablated by genetic knockout have elevated contractility and increased rate of relaxation.
Protein kinase A (PKA) Phosphorylates Troponin I which reduces its affinity for Ca++ . That also causes the muscle to relax more quickly (lucitrophic effect).
Muscarinic and adenosine receptors on the myocyte couple to inhibitory G protein and act to inhibit adenylyl cyclase and lower contractility.
Calcineurin blockade blocks that pathway. Persistent elevation of cytosolic Ca++ causes hypertrophy through activation of the growth factor Nuclear Factor of Activated T Cells (NFAT). CREB=cAMP Response Element Binding protein
Isotonic contraction is where the tension is held constant and the muscle is allowed to shorten The greater the load the slower it shortens If the length is increased at any load the speed of shortening increases The force-velocity curve can be extrapolated to zero force to determine the maximum velocity of shortening,Vmax. Notice that Vmax is the same at all lengths
Ed Sonnenblick found that changes in contractility shift the entire curve including the y axis intercept, Vmax Vmax is a length-independent index of contractility. Unfortunately Vmax is hard to measure in humans
d lnP dt pressure At the onset of the isovolumetric period of contraction fiber velocity is near Vmax because load is low. Vmax Ventricular Pressure Pressure Time Grossman et al Circ Res 1971
d lnP dt pressure The index tends to reflect the fastest fiber in the heart (bad for regional lesions) Vmax Estimate is made before the heart is fully activated. Not very sensitive to most clinical conditions
There is a tendency to treat length tension diagrams as specific for isometric and and force-velocity plots as specific for isotonic. Actually they are just two sides of the same coin.
4. Contraction 100 gm 3. At this point the weight begins stops here weight 125 to rise and the contraction becomes isotonic 100 2. Isometric contraction as ) tension builds gms 75 distance 1. The weight is lifted Weight ( 50 on the floor and Pressure (mmHg) the muscle is relaxed 25 20 40 60 80 100 Length (mm) Left ventricular volume (ml) The heavier the weight the less distance it will be lifted How far will the muscle lift the weight? For that calculation we use the length-tension diagram It applies to hearts too