Download
1 / 18
190 likes | 363 Views
Terminal deoxynucleotidyl transferase dUTP nick end labeling. Dependency on Synaptic T arget.

E N D
FIGURE 7.15 The high-affinity NGF receptor was discovered through a series of disparate observations. A. NGF was found to elicit protein phosphorylation in PC12 cells. B. The oncogene in a cancer cell line was found to be a transmembrane receptor tyrosine kinase (RTK). C. The messenger RNA for this RTK was found in extremely high levels in DRG neurons. D. When this RTK, called p140, was eliminated from PC12 cells, they became unresponsive to NGF. (Adapted from Maher, 1988; Martin-Zanca et al., 1986; MartinZanca et al., 1990; Loeb et al., 1991)
FIGURE 7.16 The NGF signal can be transduced at the tips of growing neuronal processes. Sympathetic neurons were placed in a special tissue culture system that permitted the cell bodies and neuritesto be bathed in different media. (Left) Most neurons died when grown in the absence of NGF for 30 hours. (Right) Neurons couldbekept alive by adding NGF only to the compartments with growingneurites. In both cases, an antibody against NGF was added tothe central compartment to prevent activation of TrkA. (Adapted fromCampenot, 1977, 1982; MacInnis and Campenot, 2002)
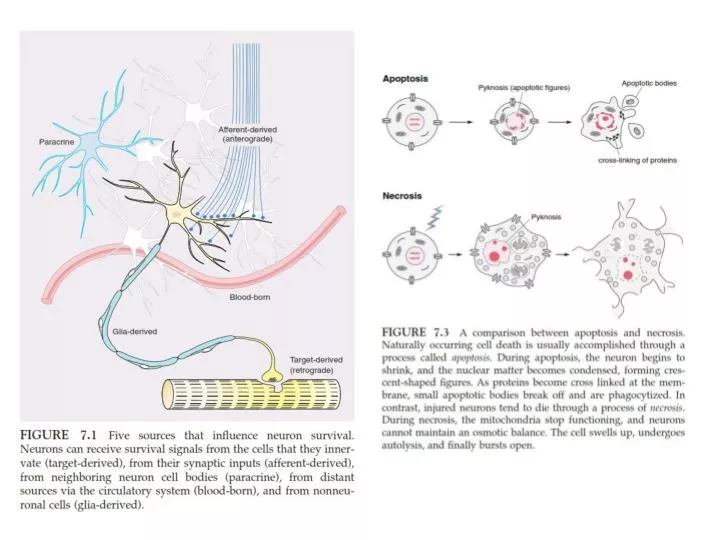
FIGURE 7.27 The molecular state of a neuron that permits it to survive or leads it to death. (Left) The living cell contains mitochondria that are preserved in a nonpermeable state due to the presence of an antiapoptoticregulator, Bcl-2. A second anti-apoptotic regulator, Bcl-x, complexes with Apaf-1, preventing the activation of caspase-9. Bcl-x also binds to a pro-apoptotic regulator, Bax, and prevents it from influencingthe mitochondrion. A different pro-apoptotic regulator, Bad, is inactive, having been phosphoryated by neurotrophin-elicited kinase activity. IAPbinds to pro-caspase-9, and this also serves to prevent activation. A nuclease that is responsible for DNA fragmentation, caspase-activated deoxyribonuclease (CAD), is bound by its inhibitor, ICAD. (Right) In dying neurons, Bad becomes active when it is dephosphorylated, and it binds to Bcl-x. This permits Bax to associate with the mitochondrion, leading to the release of cytochrome c, AIF, and Smac. It also permits Apaf-1 to form an apoptosome with cyt c, process caspase-9, and activate caspase-3. Smac binds to IAP, which also permits the processing of pro-caspase-9. One target of caspase-3 isICAD, leading to the release of CAD, and the fragmentation of DNA. In a caspase-independent pathway, AIF also enters the nucleus and fragments DNA.
FIGURE 7.29 Blocking synaptic transmission prevents normal motor neuron cell death. A. Neuromuscular transmission can be blocked by applying curare onto the chorioallantoic membrane of chick embryos. B. In control animals, over 30% of motor neurons die after embryonic day 5. When animals are treated with curare from E6-9, the magnitude of normal cell death is greatly diminished. (Panel B adopted from Pittman and Oppenheim, 1979) FIGURE 7.30 Electrical activity enhances the survival of embryonic cortex neurons by way of a neurotrophic signal. When the cultures aredepolarized by adding KCl to the culture media, calcium entersthe neurons, and the level of BDNF expression increases, leadingto greater neuron survival compared to control media. The trophicinfluence of depolarization is eliminated by adding a function-blocking anti-BDNF antibody to the growth medium. (Adopted From Ghosh et al., 1994)
FIGURE 7.31 Afferent innervation regulates neuron survival in a chick central auditory nucleus. A. Auditory neurons from the cochlea innervate the nucleus magnocellularis (NM) in the chick auditory brain stem. The removal of a cochlea (right) completely denervates NM neurons on the ipsilateral side. B. When a cochlea is removed at embryonic day 2 (E2), about 30% of NM neurons are lost during the ensuing two weeks, although cell death does not begin until E10. When the cochlea is removed at posthatch day 14, about 25% of neurons die within two days. In adults, cochlear ablation results in the loss of only about 5% of NM neurons. (Adapted from Parks, 1979; Born and Rubel, 1985)