Download

1 / 36
420 likes | 1.24k Views
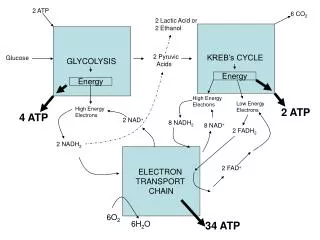
Electron transport chain-2. Introduction. The primary function of the citric acid cycle was identified as the generation of NADH and FADH2 by the oxidation of acetyl CoA. In oxidative phosphorylation, NADH and FADH2 are used to reduce molecular oxygen to water.

E N D
Introduction • The primary function of the citric acid cycle was identified as the generation of NADH and FADH2 by the oxidation of acetyl CoA. • In oxidative phosphorylation, NADH and FADH2 are used to reduce molecular oxygen to water. • The highly exergonic reduction of molecular oxygen by NADH and FADH2 occurs in a number of electrontransfer reactions, taking place in a set of membrane proteins known as the electron-transport chain.
oxidation-reduction potential • High-energy electrons and redox potentials are of fundamental importance in oxidative phosphorylation. • In oxidative phosphorylation, the electron transfer potential of NADH or FADH2 is converted into the phosphoryl transfer potential of ATP. • The measure of phosphoryl transfer potential is already familiar to us: it is given by G°´ for the hydrolysis of the activated phosphate compound. • The corresponding expression for the electron transfer potential is E´0, the reduction potential (also called the redox potential or oxidation-reduction potential).
A negative reduction potential means that the reduced form of a substance has lower affinity for electrons than does H2. • A positive reduction potential means that the reduced form of a substance has higher affinity for electrons than does H2. • Thus, a strong reducing agent (such as NADH) is poised to donate electrons and has a negative reduction potential, whereas a strong oxidizing agent (such as O2 ) is ready to accept electrons and has a positive reduction potential.
Electrons tend to pass from the most negative carrier to the most positive carrier (oxygen). This help stepwise flow of electrons.” • The standard free-energy change G°´ is related to the change in reduction potential E´° by: Δ G°´= - n f Δ E°´ Where: ΔG°´ = standard free energy n = number of electrons F = is Faraday constant (23.04 cal/volt) ΔE°´ = the difference in the standard reduction potentials and its in volt.
E´º in volts is measured by a responsive electrode placed in solution containing both the electron donor and its conjugate electron acceptor at standard conditions. 1 M concentration, 25ºC and pH 7
Example • The free-energy change of an oxidation-reduction reaction can be readily calculated from the reduction potentials of the reactants. For example, consider the reduction of pyruvate by NADH, catalyzed by lactate dehydrogenase. The reduction potential of the NAD+:NADH couple, or half-reaction, is -0.32 V, whereas that of the pyruvate: lactate couple is -0.19 V.
To obtain reaction a from reactions b and c, we need to reverse the direction of reaction c so that NADH appears on the left side of the arrow. In doing so, the sign of E´0 must be changed. For reaction b, the free energy can be calculated with n = 2. Likewise, for reaction d, Thus, the free energy for reaction a is given by
Redox potential under non-standard conditions (Nernst equation) • Under standard conditions: Δ G°= -n f Δ E° • If we not operating under standard conditions we know that Δ G = Δ G° + RT ln Keq Since : Δ G = - n f E Δ G°= -n f E° These can be combined to give -n f E = -n f E° +RT ln Keq
E = E° - RT / nf ln [ oxidant]/ [reductant] Or E = E° - RT/nf 2.303 log [ oxidant]/ [reductant] Nernst equation is used to calculate redox potential E, at any concentration of oxidant and reductant from Eº When a system is at equilibrium, ∆E = 0. We have: ∆E◦ = RT/nflnKeq Thus, the equilibrium constant and ∆E° are related
The transfer of electrons down the respiratory chain is energetically spontaneous because: - NADH is a strong electron donor - Oxygen is strong electron acceptor
How ATP becomes synthesized during the transfer of electrons to oxygen
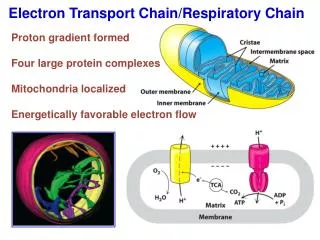
The Components of the Electron Transport Chain • The electron transport chain of the mitochondria is the means by which electrons are removed from the reduced carrier NADH and transferred to oxygen to yield H2O. • Electrons move along the electron transport chain going from donor to acceptor until they reach oxygen the ultimate electron acceptor. • The standard reduction potentials of the electron carriers are between the NADH/NAD+ couple (-0.315V) and the oxygen/H2O couple (0.816V).
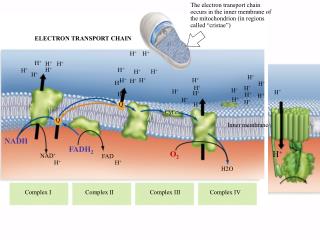
Overview of the Electron Transport Chain • The components of the electron transport chain are organized into 4 complexes. Each complex contains several different electron carriers. 1. Complex I also known as the NADH-coenzyme Q reductase or NADH dehydrogenase. 2. Complex II also known as succinate-coenzyme Q reductase or succinate dehydrogenase. 3. Complex III also known as coenzyme Q reductase. 4. Complex IV also known as cytochrome c reductase. • Each of these complexes are large multisubunit complexes embedded in the inner mitochondrial membrane.
Complex I: • Also called NADH-Coenzyme Q reductase because this large protein complex transfers 2 electrons from NADH to coenzyme Q. Complex I was known as NADH dehydrogenase. • Complex I (850,000 kD) contains a FMN prosthetic group which is absolutely required for activity and seven or more Fe-S clusters. • This complex binds NADH, transfers two electrons in the form of a hydride to FMN to produce NAD+ and FMNH2. • The subsequent steps involve the transfer of electrons one at a time to a series of iron-sulfer complexes.
The importance of FMN. First it functions as a 2 electron acceptor in the hydride transfer from NADH. Second it functions as a 1 electron donor to the series of iron sulfur clusters. • The process of transferring electrons from NADH to CoQ by complex I results in the net transport of protons from the matrix side of the inner mitochondrial membrane to the inter membrane space where the H+ ions accumulate generating a proton motive force. • The stiochiometry is 4 H+ transported per 2 electrons. NADH + H+ + CoQ NAD+ + CoQH2 ΔEo’ = 0.060 V – (−0.315V) = 0.375 V ΔGo’ = −nFΔEo’ = −72.4 kJ/mol
Complex II: • It is none other than succinate dehydrogenase, the only enzyme of the citric acid cycle that is an integral membrane protein, so its the only membrane-bound enzyme in the citric acid cycle • This complex is composed of four subunits. Two of which are iron-sulfur proteins and the other two subunits together bind FAD through a covalent link to a histidine residue.
In the first step of this complex, succinate is bound and a hydride is transferred to FAD to generate FADH2 and fumarate. • FADH2 then transfers its electrons one at a time to the Fe-S centers. Thus once again FAD functions as 2 electron acceptor and a 1 electron donor. The final step of this complex is the transfer of 2 electrons one at a time to coenzyme Q to produce CoQH2.
The overall reaction for this complex is: Succinate + CoQ Fumarate + CoQH2 ΔEo’ = 0.060 V – (+0.031V) = 0.029 V ΔGo’ = −nFΔEo’ = −5.6 kJ/mol. • For complex II the standard free energy change of the overall reaction is too small to drive the transport of protons across the inner mitochondrial membrane. • This accounts for the 1.5 ATP’s generated per FADH2 compared with the 2.5 ATP’s generated per NADH.
Complex III: • This complex is also known as coenzyme Q-cytochrome c reductase because it passes the electrons formCoQH2 to cyt c through a very unique electron transport pathway called the Q-cycle. • In complex III we find two b-type cytochromes and one c-typecytochrome.
Q-cycle: • The Q-cycle is initiated when CoQH2 diffuses through the bilipid layer to the CoQH2 binding site whichis near the intermembrane face. This CoQH2 binding site is called the QPsite. • The electron transfer occursin two steps. First one electron from CoQH2 is transferred to the Fe-S protein whichtransfers the electron to cytochrome c1. This process releases 2 protons to the intermembrane space. First half of Q-cycle
The second electron is transferred to the bLheme which converts CoQH●- to CoQ. This re-oxidized CoQ can now diffuse away from the QP binding site. The bLheme is near the P-face. The bLheme transfers its electron to the bHheme which is near the N-face. This electron is then transferred to second molecule of CoQ bound at a second CoQ binding site which is near the N-face and is called the QN binding site. This electron transfer generates a CoQ ● - radical which remains firmly bound to the QN binding site. This completes the first half of the Q cycle. First half of Q-cycle
continue • The second half of the Q-cycle is similar to the first half. A second molecule of CoQH2 binds to the QP site. In the next step, one electron from CoQH2 (bound at QP) is transferred to the Rieske protein which transfers it to cytochrome c1. This process releases another 2 protons to the intermembrane space. second half of Q-cycle
The second electron is transferred to the bLheme to generate a second molecule of re-oxidized CoQ. The bLheme transfers its electron to the bHheme. This electron is then transferred to the CoQ- radical still firmly bound to the QN binding site. The take up of two protons from the N-face produces CoQH2 which diffuses from the QN binding site. This completes Q cycle. Second half of Q-cycle
The net equation for the redox reactions of this Q cycle is: • QH2+ 2 cyt c1(oxidized) + 2H+ Q +2 cyt c1(reduced) +4H+ • Cytochrome c is a soluble protein of the intermembrane space. After its single heme accepts an electron from Complex III, cytochrome c moves to Complex IV to donate the electron
Complex IV: • Complex IV is also known as cytochrome c oxidase because it accepts the electrons from cytochrome c and directs them towards the four electron reduction of O2 to form 2 molecules of H2O. 4 cyt c (Fe2+) + 4 H+ + O2 4 cyt c (Fe3+) + 2H2O • Cytochrome c oxidase contains 2 heme centres, cytochrome a and cytochrome a3 and two copper proteins. • The reduction of oxygen involves the transfer of four electrons. Four protons are abstracted from the matrix and two protons are released into the intermembrane space.
ATP synthetase: ATPase (Complex V) • This enzyme complex synthesizes ATP , utilizing the energy of the proton gradient (proton motive force) generated by the electron transport chain. • The Chemiosmotic theory proposes that after proton have transferred to the cytosolic side of inner mitochondrial membrane, they can re-enter the matrix by passing through the proton channel in the ATPase (F0), resulting in the synthesis of ATP in (F1) subunit.
Coenzyme Q exists in mitochondria in the oxidized quinone form under aerobic conditions and in the reduced quinol form under anaerobic conditions. • Structure is similar to vitamin K and E. • All are characterized by the presence of polyisoprenoid side chain: [ CH2-CH=C-CH3-CH2]n.
The ETC contains excess of Coenzyme Q. This is compatible with Q acting as a mobile components of the ETC that collects reducing equivalents from the more fixed flavoprotein complexes and pass them to cytochromes.
Cytochromes • These are iron- containing electron transferring proteins. • They are heme proteins. • 3 classes have been identified a,b and c • Each cytochrome molecule in its ferric (Fe 3+) form accepts one electron and reduced to the ferrous state (Fe2+). • In addition to iron, Cyt a3also contain 2 bound copper atom which undergo cupric (Cu 2+) to cuprous (Cu+) redox changes during electron transfer.
Uncouplers • Electron transport and phosphorylation can be uncoupled by compounds that increase the permeability of the innermitochondrial membrane to protons in any place. i.e Uncouplers causes electron transport to be proceed at a rapid rate without the establishing of proton gradient e.g : 2,4 dinitrophenol
The energy produced by the transport of electrons is released as heat rather than being used to synthesize ATP. • In high doses, the drug aspirin uncouple oxidative phosphorylation. This explain the fever that accompanies toxic overdoses of these drugs.
Electron transport inhibitors • These compounds prevent the passage of electrons by binding to chain components, blocking the oxidation/reduction reaction • Inhibition of electron transport also inhibits ATP synthesis. e .g: - Amytal and Rotenone block e- transport between FMN and Co Q. - Antimycin A blocks between Cyt b and Cyt c - Sodium azide blocks between Cyt a a3 and oxygen
Ionophores • Ionophores are termed because of their ability to form complex with certain cations and facilitate their transport across the mitochondrial membrane. So ionophores are lipophilic e. g: Valinomycin: allows penetration of K+ across the mitochondrial membrane and then discharges the membrane potential between outside and the inside ( i.e: does not affect the pH potential). Nigericin: also acts as ionophore for K+ but in exchange with H+. It therefore abolishes the pH gradient.