Download

1 / 76
760 likes | 769 Views
Multiscale Models of Selectivity of Calcium Channels. crucial to the Physiology of Nerve and Muscle. Mathematics describes only a tiny part of life, But Mathematics* Creates our Standard of Living *e.g., electricity, computers, fluid dynamics, optics, structural mechanics, ….

E N D
Multiscale Models of Selectivity of Calcium Channels crucial to the Physiology of Nerve and Muscle
Mathematics describes only a tiny part of life, But Mathematics* Creates our Standard of Living *e.g.,electricity, computers, fluid dynamics, optics, structural mechanics, ….
How can we use mathematics to describe biological systems? I believe Some biology can be described by Physics and Mathematics “as usual”
From StructureANATOMY to Function PHYSIOLOGY using fundamental physical laws PHYSIOLOGY
Multiscale MATHEMATICAL Analysishas rarely been possible until nowCan be done for Caclium Channels
More Background Michael Fill, Ph.D., Dept. Physiology, Rush University
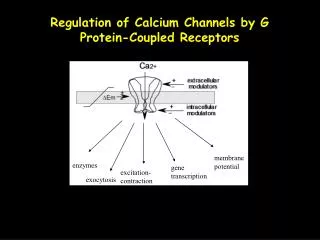
Cardiac Coupling Ca2+ DHPR-2 L type Ca Channel Ca2+ influx Ryanodine Receptor RyR-2 Ca2+ CSQ-2 Ca2+ Michael Fill, Ph.D., Dept. Physiology, Rush University L-typeCalcium Channel Ryanodine Receptor
OUTSIDE L-type Ca channel INSIDE Ca Ca RyR RyR Ca SR Calcium channels in muscle • Ryanodine receptor • large Ca2+ currentto initiate muscle contraction • Ca2+ selectivity at 10-3 M (0.1 M Na+) • DDDD locus • L-type Ca2+ channel • small Ca2+ current to change voltage or initiate Ca2+-induced Ca2+ release • Ca2+ selectivity at 10-6M (0.1 M Na+) • EEEE locus Gillespie et al., J. Phys. Chem. 109, 15598 (2005).
Ion Channels are the Main Molecular Controllers“Valves”ofBiological Function Flow timescale is 0.1 msec to min ~30 Å Figure by Raimund Dutzler
Cells and Channels Channels control flow in and out of cells ~5 µm Flow time scale is 0.1 msec to min
Ion Channels are • Biomolecular Valves, Stochastic Sensing … Hagan Bayley • Platform for Multi-Scale Technology • Simple enough to allow physical analysis of biological function • Controllers of many biological functions in health and disease • Substantial fraction of all drugs work on channels • Thousands scientists work on Ion Channels every day Thousands of scientists study channels every day
Ion Channels are Controllers of Biological Function • All signalling in the nervous system • Coordination of muscle contraction, including heart. Heart = Unintelligent Design …Denis Noble • Regulation of secretion and transport • Control of protein movement inside cells • One transporter for every molecule of special interest • Channelopathies … Fran Ashcroft Thousands of scientists study channels every day
Ion Channels are Multiscale DevicesAtomic Control of Macroscopic FlowsIon Channels are Engineering Devices
Channels control flow of Charged Spheres Channels have Simple Invariant Structure on the biological time scale. Why can’t we predict the movement of Charged Spheres through a Hole? Physics of Ionic Movement under biological conditions is MUCH SIMPLER than the physics of Fluids, Plasmas, or Semiconductors, Fluids, plasmas, and semiconductors can be computed more or less perfectly. What is the Device Equation of a Channel? Ion Channels are Devices
Where do we start? Trajectories of Charges in an Electric Field Trajectories allow Boundary Conditions and Theory of Stochastic Processes
James Clerk Maxwell “I only count molecules …., avoiding all personal enquiries which would only get me into trouble.” Slightly rephrased from Royal Society of London, 1879, Archives no. 188 in Maxwell on Heat and Statistical Mechanics, Garber, Brush and Everitt, 1995
Looking at a few trajectories in the mind’s eye (structural biology) or in simulations (molecular dynamics) is not enough. Statistical Mechanics of Devices is Required Not general nonequilibrium stat mech, Devices do not explode Temperature is well defined Distributions of velocities are Displaced Maxwellians
We start with Langevin equations of charged particles Simplest stochastic trajectories are Brownian Motion of Charged Particles Gouy-Chapman, (nonlinear) Poisson-Boltzmann, Debye-Hückel, are models with similar resolutionbut constrained to equilibrium, i.e., zero flux of all species.Devices do not exist at equilibrium. Once we learn to count Trajectories of Brownian Motion, we can count trajectories of Molecular Dynamics
Langevin Equations Positivecation, e.g., p= Na+ Negativeanion, e.g., n= Cl¯ Electric Forcefrom all charges including Permanent charge of Protein, Dielectric Boundary charges,Boundary condition charge Poisson Equation(s) Excess ‘Chemical’ Force Schuss, Nadler, Eisenberg
From Trajectories to Probabilities Main Result: Theory of Stochastic Processes Joint probability density of position and velocity satisfies a Fokker Planck equation with Fokker Planck Operator Coordinates are positions and velocities of N particles in 12N dimensional phase space Schuss, Nadler, and Eisenberg
Theory of Stochastic Processes StatisticalMechanics Device Equation Thermodynamics Schuss, Nadler, and Eisenberg
Conditional PNP Electric Force depends on Conditional Density of Charge Closures or ApproximationsNeeded ChannelProtein Nernst-Planck gives UNconditional Density of Charge mass friction Schuss, Nadler, Eisenberg
Counting at low resolution gives ‘Semiconductor Eqns’ Poisson-Nernst-Planck (PNP) Only contains correlations of means Gouy-Chapman, (nonlinear) Poisson-Boltzmann, Debye-Hückel, are siblings with similar resolutionbut without current or flux of any speciesDevices do not exist at equilibrium
PNP in 3D Poisson’s Equation Drift-diffusion & Continuity Equation Chemical Potential
Solving semiconductor equations requires a trick Poisson Equationand Nernst Planck Equation(Fick’s Law for charged particles)are solved together by Gummel iterationPoisson Transport Poisson Transport Or much better (but much harder) Newton Iteration
How do we check the theory? Compare with Biological Function! Our task is to Discover & Understand, Control & Improve Biological Function Inverse Problem
Biological function of most channels is to allow andControl Flux of Ions Our task is to Discover, Understand, Control and Improve that flux Ion Channels are Biological Devices
ExperimentTwo Synthetic Calcium ChannelsDesigned by Theory Calcium selective Unselective Wild Type As charge density increases, channel becomes calcium selectiveErev ECa built by Henk Miedema, Wim Meijberg of BioMade Corp.,Groningen, Netherlands Miedema et al, Biophys J 87: 3137–3147 (2004)
OUTSIDE L-type Ca channel INSIDE Ca Ca RyR RyR Ca SR Calcium channels in muscle • Ryanodine receptor • large Ca2+ currentto initiate muscle contraction • Ca2+ selectivity at 10-3 M (0.1 M Na+) • DDDD locus • L-type Ca2+ channel • small Ca2+ current to change voltage or initiate Ca2+-induced Ca2+ release • Ca2+ selectivity at 10-6M (0.1 M Na+) • EEEE locus Gillespie et al., J. Phys. Chem. 109, 15598 (2005).
Electrodiffusion of charged, hard spheres Putting in Correlations by Hand
Modeling Ion Flux The flux density Ji of ion species i is given by the constitutive relationship whereDiis the diffusion coefficient iis the number density iis the total chemical potential We use equilibrium Density Functional Theory of fluids to calculate the chemical potential.
Components of Chemical potential Concentration-independent Excess chemical potential Physical components (correlations, e.g., size) Chemical components (orbital delocalization) Ideal term chemical potential of uncharged point particles
The Excess Chemical Potential • Density Functional Theory* is the state-of-the-art in theories of inhomogeneous fluids (including electrolytes). • DFT computes thermodynamic quantities of confined fluids in equilibrium (e.g., average concentration profiles, surface tension). • electrolytes, colloids, polymers • one of a few theories used for inhomogeneous fluids • 100’s of papers a year use DFT of fluids. • DFT is ideal for channels because selectivity filters confine ions in a small geometry. *Fundamental measure theory of Rosenfeld, … then Roth
The Excess Chemical Potential Density Functional Theory minimizes a free energy functional Ω that depends only* on all the averageion concentration profiles i(x): How to express the Helmholtz energy of the system in terms of the averageion concentration profiles is not obvious or simple and no one knows if it is possible in general. It is important to include all correlations. *possible (approximately) for charged, hard spheres Gillespie et al
Hard-Sphere Component excluded volume: centers of two hard spheres of radius R cannot come closer than 2R Screening Component describes the electrostatic correlations of finite-size ions beyond mean-field component Components of Excess Chemical Potential radius of species k Mean-Field Component Poisson equation
Application to the Ryanodine Receptor D. Gillespie et al., J. Phys. Chem. 109, 15598 (2005).
The Ryanodine Receptor Calcium Channel • Physiological role: conduct large Ca2+ current and start contraction of muscle • RyR has the signature of a classic calcium channel • Selectivity controlled by DDDD (4 negative charges) • EEEE locus is also important Y. Wang et al., Biophys. J. 89, 256 (2005).
The Geometry L. Xu et al., Biophys. J. 90, 443-453 (2006)
The Geometry • Selectivity Filter • is 10 Å long and 8 Å in diameter • confines the four D4899 residues. • The four E4900 residues are confined to the lumenal side, overlapping D4899. • Cytosolic distributed charge Protein Protein D. Gillespie et al., J. Phys. Chem. 109, 15598 (2005).
Our Approach • Use a few select data points to compute input parameters. • channel structure (e.g., E4900 location) • ion diffusion coefficients in selectivity filter • Let the theory predict all the other data points. • >50 data curves, growing all the time thanks to LeXu! D. Gillespie et al., J. Phys. Chem. 109, 15598 (2005).
1000 | 100 mM 250 | 100 mM 1000 | 500 mM 250 | 500 mM 1000 | 1000 mM 250 | 1000 mM KCl cytosolic | lumenal concentrations D. Gillespie et al., J. Phys. Chem. 109, 15598 (2005).
KCl 250 mM cytosolic (25-2000 mM) lumenal concentrations • experiments • theory relative % error D. Gillespie et al., J. Phys. Chem. 109, 15598 (2005).
LiCl Misfit in unequal concentrations D. Gillespie et al., J. Phys. Chem. 109, 15598 (2005).
KCl CaCl2 KCl MgCl2 Divalents 250 mM | 250 mM monovalent 4 μM | ■ 5 mM divalent ■ 10 mM ■ 50 mM bi-ionic: ▲250 mM monovalent | 25 mM divalent D. Gillespie et al., J. Phys. Chem. 109, 15598 (2005).
Divalents NaCl CaCl2 CsCl CaCl2 Misfit in unequal concentrations 250 mM | 250 mM monovalent 4 μM | ■ 5 mM divalent ■ 10 mM ■ 50 mM bi-ionic: ▲250 mM monovalent | 25 mM divalent D. Gillespie et al., J. Phys. Chem. 109, 15598 (2005).
Bi-ionic Solutions KCl | LiCl 250 mM | 250 mM KCl | NaCl KCl | RbCl KCl | CsCl D. Gillespie et al., J. Phys. Chem. 109, 15598 (2005).
Mutations: Change the Charge KCl with 10 mM CaCl2 KCl only D4899N Protein charge density13 0 M Water is 55 M Gillespie et alJ Phys Chem 109 15598 (2005)
Mole Fraction Experiments D. Gillespie et al., J. Phys. Chem. 109, 15598 (2005).
Mole Fraction Experiments The model predictedanomalous mole fraction effect (AMFE) for mixtures of Na+/Cs+No AMFE for other mixtures (e.g., Li+/K+). Model predicted that AMFE disappears when 50 mM LiCl is added to the baths. It Does! D. Gillespie et al., J. Phys. Chem. 109, 15598 (2005).
Selectivity • Selectivity and permeation stem from two competing phenomena: • the charge density of the protein attracts counter-ions to high densities (10-20 M!) • Protein charges and permeantions compete for space Electrostaticscontrols (i.e., buffers) Channel Contents Forcing Crowded Chargeand thus Selectivity W. Nonner et al., Biophys. J. 79, 1976 (2000).