Download

1 / 36
370 likes | 518 Views
Co-Evolution of the Genetic Code and Amino Acid BioSynthesis. An hypothesis from 1975 by Jeffrey Tze-Fei Wong. Anna Battenhouse. Universal Phylogenetic Tree. Translation – the Players. Ribosome large subunit: 23S rRNA, many proteins peptidyl transferase reaction, tRNA sites

E N D
Co-Evolution ofthe Genetic Code andAmino Acid BioSynthesis An hypothesis from 1975 by Jeffrey Tze-Fei Wong Anna Battenhouse
Translation – the Players Ribosome • large subunit: 23S rRNA, many proteins • peptidyl transferase reaction,tRNA sites • small subunit: 16S rRNA, many proteins • messenger RNA (mRNA) contacts Translation factors • EF-Tu, EF-G proteins, GTP tRNA (transfer RNA) • acceptor arm holds amino acid • anticodon arm “reads” mRNA, implements Genetic Code aaRS (aminoacyl tRNA synthetase) • “charge” tRNAs with the appropriate amino acid 22 “coded” amino acids
DNA excellent information storage,poor catalysis transcription synthesis,metabolism poor information storage,excellent catalysis adequate information storage,adequate catalysis RNA protein translation Chicken or Egg?
Simplifying Assumptions • Ribosome proteins serve as scaffold • Small PTC RNA core with 2-fold symmetry • A, P sites • Translation factors not required • EF-Tu, EF-G, GTP • “Proto-genes” were RNA molecules • copied by an RNA replicase ribozyme • tRNA charging enzymes were ribozymes • left imprint on modern aaRSs
The Pre-translation RNA world was metabolically complex Diverse RNA enzymes (ribozymes), using cofactors and small random peptides Benner, S.A., Ellington, A.D., Tauer, A., Modern metabolism as a palimpset of the RNA world PNAS 86 (1989)
What drove code evolution? • Sterochemical interactions • Codon assignments arose from Physical/chemical interactions between AAs and RNA • Error minimization • Adjacency of codons minimizes potential damage due to mutations/translation errors • Expanding codons • Not all codon triplets used at first. Usage expanded over time to modern 64. • Amino acid biosynthesis • Formation/extension of AA biosynthetic pathways
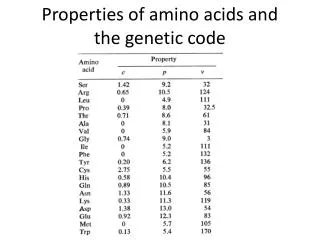
PNAS 55 (1966) 7.5 9.1 Woese et al., Microbio. Mol. Bio. Rev., 64:1 (2000)
Yarus 2009 Results • RNA can bind wide variety of AAs specifically • polar, charged, aromatic • even aliphatic • Several AA/RNA binding sites showed anticodon enrichment • Ile, Phe, Arg, His, Trp, (Tyr) • However ~80% of triplets not found 7.5 9.1 Woese, PNAS 55 (1966)
Wong, J.T., PNAS 73 (1976) Amino Acid Biosynthesis Co-Evolution Wong, J.T., Trends Bio. Sci.,Feb. 1981
BioSynthesis Co-Evo Predictions • AA biosynthesis is essential • phase 1 AA abundancy • phase 2 AA non-abundancy • Biosynthetic evolutionary trace should still be discernable for precursor product pairs • codon allocation • “pre-translation” synthesis • Set of encoded AAs is, in theory, (slightly) mutable
Cys, Met, Trp, Phe, His UV labile Asn, Gln thermally unstable Gly, Ala, Val,Leu Ile,Ser,Asp,Glu, initially most abundant Not all amino acids would initially be available/abundant
Genetic Code by Biosynthetic Families Wong, J.T., Coevolution theory of the genetic code at age 30, BioEssays, 27.4 (2005)
Indirect Charging (“pre-translation” biosynthesis) Precursor Product inventivebiosynthesis AA AA AA AA AA AA AA Amino Acyl tRNA Synthetases (“aaRSs”) tRNA charging enzymes Direct Charging
Archaea Archaea O’Donoghue et al.,PNAS 102:52 (2005) Sep-tRNA Cys-tRNA(Sep = O-phosphoserine) Lack of CysRS Euryarchaea Pre-translation Biosynthesis Wong, J.T.,BioEssays 27.4 (2005)
neither precursor nor product aaRS precursor aaRS only both precursor and product aaRS Distribution of Genes forPre-trans biosynthesis Glu Gln Asp Asn Wong, J.T., Coevolution theory of the genetic code at age 30, BioEssays 27.4 (2005)
Additional Evidence • Phylogeny of aaRS genes • product aaRSs are often related to their precursor aaRSs (and precursors more ancient) • Enzyme for de novoAsn synthesis in many archaea was once an AspRS • pre-trans de novo biosynthesis via aaRS paralog • Natural and synthetic modifications to the Genetic code exist • pyrrolysine – 22nd amino acid • engineered AA additions in E. coli Roy et al., and Francklyn, C., PNAS 100:17 (2003); Doring, et al., Scienece 292:501 (2001)
Pyrrolysine • Incorporated in only a few prokaryotic proteins • has its own tRNA, (codon UAG, normally “stop”), aaRS • Found in only a few species • Archaea • 3 Methanosarcina • Methanococcoides • Eubacteria • Desulfitobacterium hafniense (HGT) • All species live off methylamine (fishy smell) • Pyl used in monomethylamine methyltransferase enzyme Lehninger, Principles of Biochemistry, Fifth Ed.
BioSynth Co-Evo Theory Limitations • Long on correlations, short on mechanisms • Does not address the important questions surrounding tRNA • how did it arise? • did the anticodon arm develop independently of the acceptor stem? • how did aaRSs come to be? • and the Class I/Class II aaRS division • role of the extensive AA base modifications • What about the co-evolution of tRNAs and the 23S and 16S RNAs? • and the fascinating questions around message-reading translocation
Acceptor stem Anticodon wobble position Maizels, N. et al., Biol. Bull. 196 (1999) Transfer RNA (tRNA)
Class I aaRSs Class II aaRSs • Beta sheet active site • 3’ –OH attachment • interacts with major groove of tRNA acceptor stem • Rossman fold active site • 2’ –OH attachment first • interacts with minor groove of tRNA acceptor stem Schimmel et al., in The RNA World, Third Edition, Cold Spring Harbor Laboratory Press (2006)
tRNA Identity Elements Giege, R. et al., Nucleic Acids Res. 26 (1998)
Class I aaRS Class II aaRS Giege, R. et al., Nucleic Acids Res. 26 (1998)
Xue, H., Tong, K., Marck, C., Grosjean, H., Wong, J.T., Transfer RNA paralogs, Gene 310 (2003) tRNA phylogeny
Wobble I (inosine) can pair with C,U,A Watson/Crick A-U pair Non-Watson/Crick G-U pair
Tong, K., Wong, J.T., Anticodon and wobble evolution, Gene 333 (2004)