Download
1 / 34
340 likes | 782 Views
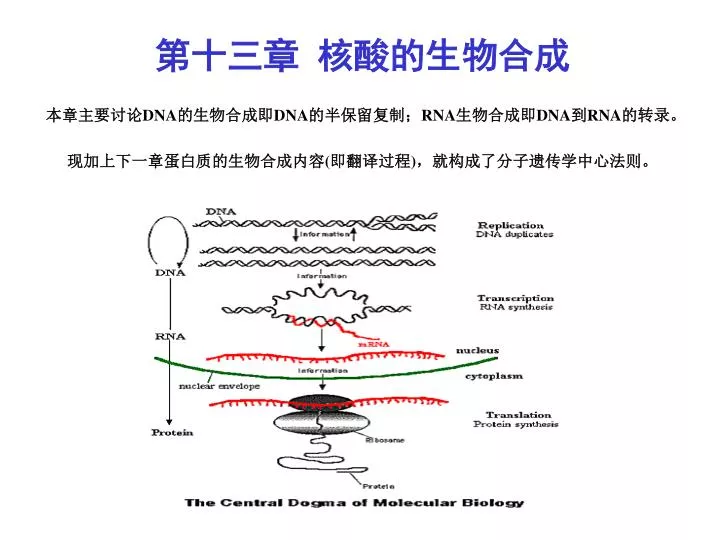
第十三章 核酸的生物合成 本章主要讨论 DNA 的生物合成即 DNA 的半保留复制; RNA 生物合成即 DNA 到 RNA 的转录。现加上下一章蛋白质的生物合成内容(即翻译过程),就构成了分子遗传学中心法则。. 第一节 DNA 的复制与修复 研究 DNA 复制的目的就是要了解三个问题。 第一、子代 DNA 为什么能够直接地获得新 DNA 的遗传信息? 第二、复制是怎样进行的 第三、生物体是怎样对 DNA 复制进行调控的? 一、 DNA 的半保留复制 ( P321 图19-1 P321 图19-1)

E N D
第十三章 核酸的生物合成本章主要讨论DNA的生物合成即DNA的半保留复制;RNA生物合成即DNA到RNA的转录。现加上下一章蛋白质的生物合成内容(即翻译过程),就构成了分子遗传学中心法则。
第一节 DNA的复制与修复 研究DNA复制的目的就是要了解三个问题。 第一、子代DNA为什么能够直接地获得新DNA的遗传信息? 第二、复制是怎样进行的 第三、生物体是怎样对DNA复制进行调控的? 一、DNA的半保留复制 (P321 图19-1 P321 图19-1) 在DNA复制过程中,每个子代分子的一条链来自亲代DNA,另一条则是新合成的,这种复制方式称为半保留复制。实验证明见课本。 生物意义:DNA的半保留复制机制可以说明DNA在代谢上的稳定性。
(一)、反应所需的条件及反应的特点: (1)原料(或底物):四种5′三磷酸脱氧核苷(dNTP),缺一不可。 (2)模板及辅助因子:DNA模板、Mg2+(或Mn2+)或DNA二条链(一条链为模板,一条链为引物) (3)引物链:具有自由3′-OH的RNA或DNA片段(约10个核苷酸) (4)反应:由引物末端3′-OH与进入的5′三磷酸脱氧核苷的α磷酸残基化合,生成酯键(3′5′磷酸二酯键,并脱下焦磷酸) (5)方向:DNA 链由5′→3′方向延长 (6)能量:来自α与β之间的高能磷酸键裂解 (7)四种dNTP参加反应的先后顺序,由DNA模板决定,受碱基配对支配;而不受四种dNTP相对浓度的影响。 (8)产物DNA的性质与模板相同。这说明DNA聚合酶是一种模板指导的酶。
二、参加复制过程的主要酶及作用 DNA是由脱氧核糖核苷酸聚合而成,参加聚合反应的酶包括多种DNA聚合酶以及DNA连接酶。 (一)大肠杆菌中DNA聚合酶:(含Zn2+)(DDDP) (1)DNA聚合物Ⅰ(又称Kornberg酶)的特殊作用:(是一个多功能酶) ①去除引物及填补间隙作用:在DNA复制过程中,可将引物RNA水解下来,并催化合成DNA片段以填补间隙。 ②DNA聚合酶作用:催化DNA链由5′→3′方向延长。 ③校正错误作用:它可将DNA链的末端接上的错误核苷酸水解下来,然后再催化接上一个正确的核苷酸。即由3′端水解DNA链,具有3′→5′核酸外切酶的作用。 ④修复损坏及变异作用:即由5′端水解DNA链,具有5′→3′核酸外切酶作用。(详见后面介绍) ⑤由3′端使DNA链发生焦磷酸解。 ⑥催化无机焦磷酸盐与脱氧核糖核苷三磷酸之间的焦磷酸基交换。
(2)DNA聚合酶Ⅱ特殊作用:目前尚不大清楚,可能在DNA修复中起作用。 (3)DNA聚合酶Ⅲ的特殊作用:主要参与绝大多数新的DNA的合成。 DNA聚合酶Ⅰ M.W. 109Kdal 单一多肽链,聚合1000个核苷酸/分/分子 DNA聚合酶Ⅱ M.W. 120Kdal 单一多肽链,聚合2000个核苷酸/分/分子 DNA聚合酶Ⅲ M.W. 180Kdal 单一多肽链,聚合50000个核苷酸/分/分子
(二)DNA连接酶及作用:DNA聚合酶能催化DNA片段及链的延长合成,但不能将DNA断片连接起来,这种连接反应是由DNA连接酶来催化的。DNA连接酶不单是DNA复制所必需的,而且也是在DNA损伤修复及基因重组中不可缺少的酶。(二)DNA连接酶及作用:DNA聚合酶能催化DNA片段及链的延长合成,但不能将DNA断片连接起来,这种连接反应是由DNA连接酶来催化的。DNA连接酶不单是DNA复制所必需的,而且也是在DNA损伤修复及基因重组中不可缺少的酶。 • 连接双股DNA中的一股存在的缺口, • 不能连接单股DNA 。
(三)拓扑异构酶: 生物体内DNA分子通常处于负超螺旋状态.
拓扑异构酶的作用 • 拓扑异构酶的作用是使DNA的超螺旋的圈数增加或者减少。 • 此作用需要ATP分解提供能量。
(五)引发酶(引物合成酶) 又称特定的RNA聚合酶 根据DNA的顺序合成RNA引物,本质上是 RNA聚合酶 RNA引物(约10个核苷酸的RAN片段)是由引 发酶(引发体)来合成的,合成时不需要引物,其碱基与一般模板DNA配对,合成的方向是5′→3′。
DNA解链后,DNA聚合酶即以分开了的两条多核苷酸链为模板进行复制。DNA两条链都能作为模板。以复制叉向前移动的方向为标准,走向为3′→5′的模板链上的DNA能以5′→3′方向连续合成,直到所需的长度,这条链能连续合成称为前导链,而另一条走向为5′→3′的模板链却不能连续合成,称为滞后链。1968年日本学者冈崎等提出了DNA的不连续复制模型,认为需先合成一些约为1000~2000个核苷酸(真核生物为100~200个)DNA片段(称为冈崎片段),其合成方向也是5′→3′,但与复制叉移动的方向相反,这些片段合成后可在DNA连接酶作用下拼接起来,直到其长度与前导链相等。DNA这一复制过程称为半不连续合成(Semidis-continous)。
四、DNA复制的主要过程:(见P342 图19-17) 1、拓扑异构酶Ⅱ可引入负超螺旋而造成DNA双链断裂,以便于解螺旋酶的作用。 2、解螺旋酶将双股螺旋解开,形成复制岔口(复制叉)。 3、单链结合蛋白质(SSB)结合于每股链上,以维持两链处于分开状态。 4、引物合成酶催化合成RNA引物。 5、DNA聚合酶Ⅲ催化合成新的DNA的前导链及冈崎片断。 6、DNA聚合酶Ⅰ切除引物,并合成DNA片段以填补空隙。 7、DNA连接酶将冈崎片断拼接起来,以完成滞后链的合成。
五、真核生物DNA的复制:复制条件、酶及因子 等均与原核生物相似。 特点 1、真核生物DNA复制的冈崎片断约为200bp,相当于一个核小体DNA的长度。(小于原核生物:1000~2000 bp) 2、复制速度比原核生物慢,基因组较大,但真核生物染色体DNA上有许多复制起点,它们可以分段进行复制。细菌的DNA复制叉移动速度为5万bp/分;哺乳动物则为1千~3千bp/分。 3、真核生物一个复制起点一般发动一次复制过程,快速生长往往采用更多的复制起点。原核生物:一个起点可连续发动复制。 4、真核生物在复制子上,由于其染色体是以核小体为结构单位组成,因此在其复制时涉及到亲代DNA链与组蛋白八聚体的解开和子代DNA与组蛋白的重新组装。
六、DNA复制调控(了解): 1963年 Jacob等提出复制子模型(即一个复制单位):(P 344) 它的一个结构基因能产生一种蛋白质——起始因子(initiator),具有 感受细胞内信号、识别复制基因、促使复制开始等作用。 复制子中存在正、负调节系统,调控机制很复杂。
七、DNA损伤及修复: (一)DNA损伤:指DNA分子的一级结构的任何异常改变,包括异常修饰和顺序改变。 可分为自发的损伤和环境因子引起的损伤。 1、物理损伤:μV辐射、电离辐射。(放射性同位素β粒子、x射线) 可使DNA分子中出现 :TT(二聚体)防碍复制、转录等。 2、化学损伤:主要有亚硝酸、亚硝基胺、甲基化试剂、碱基类似物等 改变碱基配对顺序,影响DNA正常复制。 如:亚硝酸(HNO2)可使 A变成 I (脱NH3) C变成 U (脱NH3)
(二)DNA修复:目前已知细胞内有四种修复系统:(二)DNA修复:目前已知细胞内有四种修复系统: 光复活;切除修复(复制前修复);重组修复(复制后修复);诱导修复和应急反应(SOS),后三种又称为暗修复。 修复工作相继由四种酶催化,分为四步进行:(参见P348,图19-24右半部) ①切开(特异的核酸内切酶催化):切开二聚体的5′端一侧, ②切除(核酸外切酶催化):切除含二聚体的片断。 ③修复(DNA聚合酶Ⅰ催化):在敞开的3′-OH上逐个接上正确的脱氧核苷,形成与另一链有互补关系的短链。 ④连接(DNA连接酶):将新合成的正确片段3′-OH与切去缺陷片断后而敞开的5′-P,拼接起来。 生物学意义:能使正常机体在内外环境因子高速度变化的条件下,保持遗传信息稳定和复制的准确性。
第二节 RNA的生物合成与加工 现已肯定:细胞内所有RNA都是以DNA为模板在胞核中合成,其绝大部分进入胞液中,发挥其指导蛋白质合成的功能,只有小部分留在胞核中起结构及调节作用。 RNA:分为三种:mRNA( 信使RNA)、tRNA(转运RNA)、rRNA(核糖体RNA)。 mRNA:将从DNA转录来的遗传信息传递给蛋白质,作为蛋白质合成的直接模板。一分子mRNA,可表达一个或一组基因。 tRNA:将活化了的AA运送到核蛋白体供给合成蛋白质之用。 rRNA:组成核蛋白体(核糖体),作为细胞内蛋白质合成场所,细胞内RNA的合成可由DNA指导的RNA聚合酶、RNA指导的RNA聚合酶、多核苷酸磷酸化酶等催化。但转录作用只有在DNA指导的RNA聚合酶的作用下才发生。
(一)反应所需条件及反应特点: 1、原料(或底物):四种5′-三磷酸核苷(NTP)缺一不可。 2、模板及因子:DNA模板、Mg2+或Mn2+,一般双链中只有一条链作为转录的模板链。 3、反应进入的核苷酸的5′-P与另一核苷酸的3′-OH形成磷酸二酯键。 4、反应方向:5′→3′。 5、四种NTP参加的先后顺序:由DNA模板决定,受碱基配对支配,而不受四种NTP相对浓度影响。
(二)与DNA聚合酶的不同之处 1、不需要引物; 2、作用方式不同;DNA碱基顺序的转录是通过全保留方进行的。 3、不具有核酸酶的活性(RNase或DNase的活性)
(三)RNA聚合酶的种类和组成: 1、大肠杆菌的RNA聚合酶组成: 全酶分子量约为46万;它包括两条α链(α2)、一条β链(β)、一条β′链(β′)、一分子σ因子(σ),其中σ因子的功能是识别模板上特殊起始点,使DDRP结合到DNA启动子上,故在转录开始后即被释放,剩余α2ββ′部分称为核心酶,催化转录继续进行。 2、现真核细胞中有酶Ⅰ(A)、酶Ⅱ(B)、酶Ⅲ(C)都是金属酶。(含Zn2+、Mg2+、或.Mn2+)它们的结构分布部位及功能都不完全相同 ①酶Ⅰ分布于核仁中,催化合成核仁的rRNA(18s、28s、5.8s) ②酶Ⅱ在核质中催化合成mRNA的前体及病毒RNA ③酶Ⅲ在核质中催化合成5sRNA及tRNA。 除了上述细胞核RNA聚合酶外,还分离到线粒体RNA聚合酶和叶绿RNA体聚合酶。其功能详见P362。
二、转录的基本步骤:有四个步骤(有的称三个步骤):见P361 图20-2 识别:对双链DNA特定部位的识别。 起始:聚合作用即转录的开始。 延长:RNA链的延长。 终止:聚合作用停止。 (一)识别起始: 1:识别:原核细胞RNA聚合酶的因子能辨认DNA模板链上的起始位点。(该位点被叫作启动基因或启动子)。起始点约由6—11个核苷酸构成,且具有一定的排列顺序,RNA聚合酶与起始点结合子前,DNA的两条链必须解开,具体是由于什么因子的作用尚不清楚。总之,识别可能是RNA聚合酶在一些辅助因素的协作下与DNAA链上起始点结合的步骤。 2:起始:头两个与DNA链上碱基配对的NTP依次进行聚合作用形成第一个磷酸二酯键。其中第一个核苷酸80%以上都是嘌呤,大多是GTP、(或ATP)。:DNA模板链走向:3′→5′RNA的合成方向:5′→3
(二)延长: 当RNA链合成开始后,σ因子就脱落下来,可与另一核心酶结合而被重新使用。为了能沿模板移动,全酶必须放松对DNA的结合,这些变化使得:此时RNA聚合酶构象改变:全酶→核心酶+σ因子。参见P361。核心酶可沿DNA链滑行向前,利用NTP为原料,连续合成RNA,不断延长RNA链,延伸时所合成的多核苷酸链并不全保留在模板上,对一条RNA合成链而言,在任何时候只有一个连接点,该点沿DNA链移动,连续转录出相同的RNA产物。当核心酶滑向前时,已被转录的DNA前段分开的部分链又重新并合,形成双螺旋,这一开一合逐步不断进行,直到转录过程结束。
(三)终止 在DNA分子上有终止转录的特殊信号(终止子),这些信号一些可被RNA聚合酶自身识别,另一些则需ρ因子帮助(终止辅助因子,又称终止因子)。 (1) E.Coli中存在两类终止子:①不依赖于ρ的终止子(简单终止子)。 ②依赖于ρ的终止子。 ①不依赖于ρ的终止子(简单终止子):现已弄清许多DNA分子的终止区域,发现在终止点之前有一双重对称的G与C丰富区,紧接着还有一A和T多的排列顺序。因此在些区域内转录的RNA能自身互补(G……C)而形成发夹形结构,AT丰富区有一连串的AAAA,故新生RNA链终止末端带有多个U残基(UUUU),很可能在这些结构中一个或多个即是终止信号。 ②依赖于ρ的终止子:在某些终止点有种特殊蛋白质ρ蛋白(rho protein),其分子量为5.5万,能辩认终止信号,并与酶结合,以阻止核心酶再向前滑动,于是转录作用停止,并释放出产物RNA、ρ因子、RNA聚合酶。在依赖ρ因子的终止点处,RNA聚合酶只是在此暂停,但不终止,需ρ的帮助,才能停止转录。
(2)ρ因子具有两种活性: a、终止转录; b、依赖于RNA的核苷三磷酸酶(NTPase)。因此提出了一个ρ终止转录模型。
三、转录的不对称性: 在转录过程中作为模板的DNA大多为双链的,实验证明,不管任何一个基因(DNA片断),其DNA双股链中都只有一股被转录,即所谓的不对称转录。究竟DNA双链的哪一条上带有被转录的信息呢?目前认为没有一定的约束和规定 四、一些名词: (一)启动子(promoter)和转录因子; (二) 终止子(terminator)和终止因子; (三)操纵子(operon) 详见P367转录过程的调节控制
五、RNA的转录后加工(RNA的成熟):P369 (一)RNA前体的加工处理(修饰作用)主要包括: 1、剪切:5′或3′端的切除 2、链的裂解: 3、特殊结构的形成:如mRNA 5′—帽子的形成。 4、碱基或糖的修饰:如甲基化。 5、拼接:切除内含子,拼接外显子。 (二)tTNA前体的加工: 1、切除:用核酸内切酶、核酸外切酶切除5′端和3′端的附加序列。 2、碱基修饰:特异的修饰酶催化。
3、加上3′-CCAOH结构,它对于接受氨酰基的活性是必要的,该反应是由tRNA核苷酰转移酶催化进行,由CTP和ATP供给胞苷酸和腺苷酸。成熟tRNA分子较小(MW∶2.4万~3.1万),沉降系数4s±,具有二个功能部位(3′-CCAOH即AA臂;反密码环)。tRNA种类多(50多种),而体内AA只有20种,故可有几种tRNA转运同一种AA称为同功受体tRNA,如转运苯丙氨酸的二种tRNA可用tRNAphe表示。3、加上3′-CCAOH结构,它对于接受氨酰基的活性是必要的,该反应是由tRNA核苷酰转移酶催化进行,由CTP和ATP供给胞苷酸和腺苷酸。成熟tRNA分子较小(MW∶2.4万~3.1万),沉降系数4s±,具有二个功能部位(3′-CCAOH即AA臂;反密码环)。tRNA种类多(50多种),而体内AA只有20种,故可有几种tRNA转运同一种AA称为同功受体tRNA,如转运苯丙氨酸的二种tRNA可用tRNAphe表示。 (三)rRNA的前体加工: 占细胞内RNA总量的80%,由于①rRNA和prot 结合存在,②核蛋白体在功能上须反复使用,所以代谢比较稳定。, rRNA包括28s、18s、5.8srRNA,在核仁中合成; 5srRNA在核质中合成。 转录后rRNA前体的加工过程主要包括:拼接、断裂、甲基化等反应 rRNA的功能主要是:组成核蛋白体,为prot合成的“装配机”。
(四)mRNA 占细胞总量最少,真核生物的mRNA前体是核不均一RNA(HnRNA),须经复杂的加工过程才能变成成熟mRNA,进入细胞质胞液中作为蛋白质合成模板。
hnRNA其加工过程包括: 1. 5′端“帽子”结构的形成;5′—“-帽子”→非翻译顺序→ 起始顺序(多AUG)→密码区→”尾巴”。 2 多聚A—‘尾巴”的形成; 3. 链内核苷甲基化; 4 . 除去内含子,拼接外显子。
