Download

1 / 46
550 likes | 2.08k Views
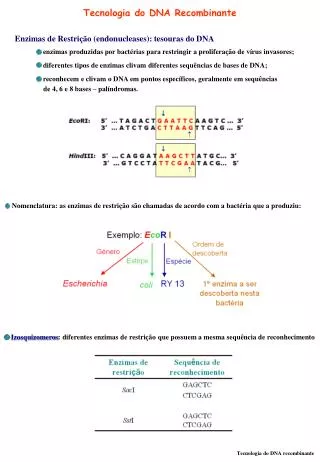
. Ha reso possibile il CLONAGGIO dei GENI permettendo di ISOLARE AMPLIFICARE frammenti di DNA SEQUENZIARE . TECNOLOGIA DEL DNA RICOMBINANTE. Perché manipolare i geni? 1) Per facilitare lo studio dell’espressione genica e della regolazione fisiologica;

E N D
Ha reso possibile il CLONAGGIO dei GENIpermettendo di ISOLARE AMPLIFICARE frammenti di DNA SEQUENZIARE TECNOLOGIA DEL DNA RICOMBINANTE Perché manipolare i geni? 1) Per facilitare lo studio dell’espressione genica e della regolazione fisiologica; 2) per identificare il prodotto di un gene e/o per ottenerne la sovraespressione; 3) per studiare la relazione fra struttura e funzione delle proteine; 4) per identificare componenti cellulari che interagiscono con particolari sequenze di acidi nucleici o con particolari domini proteici
PROCEDURA di CLONAGGIO ISOLAMENTO del gene INSERZIONE del gene in un VETTORE PLASMIDICO INTRODUZIONE del vettore plasmidico IN CELLULE VIVENTI per propagarlo Le fasi più delicate sono quelle del taglio e dell’unione di sequenze di DNA in modo preciso…. …..tutto ciò si ottiene con l’ausilio di ENZIMI
Cosa serve per un clonaggio • Gene (DNA di interesse) • Enzimi di restrizione • DNA ligasi • Vettore • Cellula ospite
CARATTERISTICHE DELLA REAZIONE: Presenza di ATP 10° C o.n. oppure temperatura ambiente, poche ore P P P P P P P P La bassa temperatura è consigliata in quanto, diminuendo l’energia cinetica delle molecole, riduce la possibilità che le estremità appaiate si separino prima di venir stabilizzate dalla ligazione. OH OH OH EcoRI BamHI OH 5’-G AATTC-3’ 3’-CTTAA G-5’ 5’-G GATCC-3’ 3’-CCTAG G-5’ OH OH OH OH DNA LIGASI Forma legami covalenti fra il gruppo fosfato 5’ di una estremità ed il gruppo ossidrilico 3’ della catena adiacente. L’enzima comunemente utilizzato è la T4 DNA Ligasiperché stabile e poco costosa.
DNA LIGASI BamHI OH OH 5’-G GATCC-3’ 3’-CCTAG G-5’ OH OH OH OH OH OH FOSFATASI ALCALINA P P P P P P P P P P P P P P P P OH OH OH OH OH OH OH OH Blunt ends ligation oppure trattamento con Transferasi terminale Sticky ends ligation HpaI OH 5’-GTT AAC-3’ 3’-CAA TTG-5’ OH AAA OH + dATP OH AAA Sticky ends ligation TTT TTT + dTTP
Vettori di clonaggio • Plasmidi • Fagi • Cosmidi • Yac • Bac
Piccole molecole di DNA batterico extracromosomico, circolare. In natura conferiscono vantaggi selettivi ai ceppi che li contengono. Ingegnerizzati per l’uso di laboratorio. Per essere un buon vettore di clonaggio, un plasmide deve avere: Dimensioni contenute Un sito di origine della replicazione Un gene per la resistenza a un antibiotico Siti unici di restrizione (polilinker) PLASMIDI
TRASFORMAZIONE delle CELLULE BATTTERICHE: Inserimento del plasmide nella cellula ospite Cellule competenti Solo le cellule che hanno incorporato il plasmide formeranno delle colonie.
INTERVALLO (Lag phase) I batteri vengono diluiti nella coltura iniziale; la divisione procede lentamente in quanto le cellule si stanno adattando al terreno fresco. 5.0 4.5 4.0 3.5 3.0 2.5 2.0 1.5 1.0 0.5 0 4 - 5 ore Densità cellulare (OD600) FASE LOGARITMICA I batteri crescono esponenzialmente. 10 - 11 ore 0 5 10 15 20 Tempo (h) FASE STAZIONARIA La densità cellulare rimane costante. La coltura può anche entrare in una fase di declino in cui le cellule si lisano ed il DNA si degrada parzialmente. Curva di crescita di E. coli
1. RISOSPENSIONE 3. NEUTRALIZZAZIONE KAc E. coli NaOH / SDS RNasi Centrifugazione 2. LISI Supernatante contenente il DNA plasmidico Precipitato contenente DNA genomico, proteine, detriti cellulari DNA cromosomico DNA plasmidico PURIFICAZIONE del DNA PLASMIDICO LISI ALCALINA
DNA cromosomico DNA plasmidico Bromuro d’etidio Bromuro d’etidio Svolgimento della doppia elica e diminuzione della densità idrodinamica Svolgimento della doppia elica compensato dall’introduzione di supereliche ...PURIFICAZIONE del DNA PLASMIDICO ULTRACENTRIFUGAZIONE in gradiente di densita’ di CsCl in presenza di EtBr Il superavvolgimento indotto nel DNA plasmidico dall’intercalazione del bromuro d’etidio fra le basi ne impedisce progressivamente il legame rendendo la densità idrodinamica del DNA plasmidico maggiore di quella del DNA cromosomico.
Topoisomerasi Tyr-274 O P OH Prodotto di PCR T OH P O Tyr-274
Libreria genomica Quando si clona l’intero genoma di un organismo si dice che si costruisce una libreria (library) genomica
Caricamento Lavaggio Eluizione TTTTT TTTTT TTTTT TTTTT TTTTT TTTTT AAAAA TTTTT TTTTT TTTTT TTTTT TTTTT TTTTT AAAAAAAA AAAAAAAA TTTTT TTTTT TTTTT TTTTT TTTTT TTTTT AAAAA TTTTT TTTTT TTTTT TTTTT TTTTT TTTTT TTTTT TTTTT TTTTT TTTTT TTTTT TTTTT TTTTT TTTTT TTTTT TTTTT TTTTT TTTTT AAAAAAAA AAAAAAAA AAAAAAAA Libreria di cDNA (DNA complementare) La miscela di frammenti che contiene tutte le sequenze codificanti costituisce la “libreria di cDNA”. 1-Estrazione di mRNA
AAAAAAA 3’ mRNA 5’ + oligo dT 5’ AAAAAAA 3’ mRNA TTTTTTT 5’ + trascriptasi inversa 5’ AAAAAAA 3’ mRNA TTTTTTT 5’ DNA 3’ Rimozione del mRNA (degradazione alcalina o calore) Formazione della forcina 3’ 3’ TTTTTTT 5’ TTTTTTT 5’ Dna polimerasi Sintesi del filamento complementare 3’ TTTTTTT 5’ 3’ AAAAAAA TTTTTTT 5’ Rimozione della forcina con la nucleasi S1 3’ AAAAAAA cDNAA TTTTTTT 5’ 2-Sintesi del cDNA (1° metodo)
genoma di 1.85 x 108 bp • dopo digestione, abbiamo avuto frammenti di circa 20.000 basi che possiamo clonare nel vettore opportuno. • Se ogni tratto del genoma fosse presente una sola volta, basterebbero 10.000 cloni per rappresentare tutto il genoma. • Esiste una formula che permette di calcolare il numero dei cloni necessari per avere un alta probabilità che tutti frammenti siano rappresentati: • n = ln(1-p)/ln(1-f) • f = 20.000 bp/185.000.000 = circa 10-4 cioè ogni inserto di 20 kb è 1/10.000 del genoma totale. • La probabilità p può essere scelta come 95%, 99%, 99.9% a seconda della sicurezza che si desidera avere. • Nei tre casi il numero dei cloni sarà: • 30.000, 4-50.000, 70.000
Placche o colonie Replica su filtri di nitrocellulosa (conservare le piastre originali) • • • • • • Denaturazione, neutralizzazione e legame del DNA al filtro Punti di riferimento per l’orientazione della replica al termine dell’esperimento • • • • Recupero delle placche positive dalla piastra originale • • • • • Sonda ibridizzata Autoradiogramma che mostra la posizione delle sonde ibridizzate Autoradiografia Ibridizzazione con sonda marcata e lavaggio della sonda non legata 32P 32P 32P 32P 32P 32P Sonda non legata SCREENING mediante IBRIDIZZAZIONE SU PLACCA
Una sonda è un filamento di DNA complementare a quello che si vuole isolare dalla biblioteca. STRATEGIE DI SINTESI: CASO 1: CASO 2: E’ nota la sequenza nucleotidica del gene di interesse E’ nota la sequenza amminoacidica della proteina codificata dal gene Si predice la sequenza nucleotidica che dovrebbe codificare per quella proteina Si sintetizzano chimicamente le sequenze nucleotidiche (degenerate) più appropriate o si amplifica un frammento del gene di interesse mediante PCR Nello screening di una libreria genica il cDNA può essere utilizzato come sonda per l’isolamento del corrispondente gene. SONDE OLIGONUCLEOTIDICHE
Altri vettori di clonaggio • Fagi • Cosmidi • Yac (yeast artificial chromosome) • BAC
I fagi (o batteriofagi) sono strutture biologiche complesse in grado di fissarsi alla parete batterica e quindi introdurre il loro DNA all'interno del batterio. (trasfezione) . Il fago più usato e conosciuto è il fago lambda. • Il fago l è un fago temperato, ha un peso molecolare di 31.106 Da ed è lungo 48.502 bp
Il DNA isolato da alcune particelle virali è a doppio strand con due estremità a singolo filamento di 12 nucletotidi complementari fra loro, chiamati siti cos. Questi siti permettono al DNA di circolarizzare dopo l’infezione della cellula ospite. • 5’ GGGCGGCGACCTCGC---------------------- ACG 3’ • GCG---------------------TCGCCCGCCGCTGG • La mappa genetica del fago l comprende circa 40 geni che possono essere suddivisi in tre gruppi funzionali: • la parte sinistra, comprendente i geni da A a J, codifica per proteine strutturali della testa e della coda. • la parte centrale, contiene geni responsabili per la lisogenia, cioé il processo che porta all'integrazione del DNA virale ed altri processi ricombinativi. Gran parte di questa regione non é essenziale per la crescita litica e può essere eliminata per la costruzione di vettori. • la parte destracontiene geni coinvolti nella replicazione del DNA e nel ciclo litico (S, R), geni regolatori (cI, cII, cIII, cro, N, Q) (Fig. 33)
CICLO LISOGENICO • Ad alto stato di infezione la cellula contiene alte concentrazioni di cIII: • Hfl è inibita • cII stimola la trascrizione di cI e la proteina Integrasi • cI è un repressore dei geni del ciclo litico e blocca la sintesi di N, O, P e Q legandosi agli operatori OL e OR, bloccando il ciclo litico • CICLO LITICO • A basse concentrazioni di cIII: • Hfl non inibita • riduce l’attività o la quantità di cII • blocco della sintesi di cI • la sintesi di N, O, P, Q non è più bloccata
IMPACCAMENTO IN VITRO Geni fagici Geni fagici Proteine di assembaggio pD Proteine di assembaggio Ceppo lisogenico di E.coli portatore della mutazione amber BHB2690. L’induzione porta alla produzione di di code di l, dei precursori delle teste e delle proteine di assemblaggio, ma non di proteina D. L’impaccamento è impedito a causa della mancnaza di proteina D Ceppo lisogenico di E.coli portatore della mutazione amber BHB2688. L’induzione porta alla produzione di di code di l, di proteina D e delle proteine di assemblaggio, ma non di proteina E. Non si formano i precursori delle teste. LISI E MISCELAZIONE DEGLI ESTRATTI + AGGIUNTA DI ATP + DNA CONCATAMENRIZZATO COS COS COS COS + + + Proteine di assembaggio pD PARTICELLA FAGICA MATURA
La regione tra i geni J e N del genoma di l non è essenziale per la crescita litica. In linea di principio, un vettore privo di questa regione potrebbe contenere circa 14.500 bp di DNA estraneo • Nella testa del fago può trovare posto fino al 105% della lunghezza del suo DNA e, inoltre, esistono nei bracci di l altre regioni non essenziali che possono essere rimosse. • Considerando tutto questo si ottiene un valore massimo di circa 22 Kb di DNA estraneo inseribile. • Esiste anche un limite inferiore, pari al 75% del genoma di l, pari a circa 37 Kb, al di sotto il DNA non viene impaccato e il l non è vitale. • Esistono due tipi di vettori l • * I vettori d'inserzione • * I vettori di sostituzione
Un vettore comunemente utilizzato è il batteriofago l, lungo 49 Kb. Eliminazione del DNA non essenziale DNA virale Regione non essenziale (circa 20 Kb) DNA ricombinante DNA da clonare Assemblaggio in vitro delle particelle virali Teste e code del virus Il DNA virale entra nelle cellule, viene replicato e dirige la sintesi delle proteine virali. La lisi delle cellule rilascia le nuove particelle virali assemblate Infezione dei batteri Recupero delle particelle virali e isolamento del DNA clonato DNA VIRALE come VETTORE DI CLONAGGIO
• Estremità coesive Sito cos Estremità coesive All’interno della cellula, le estremità si appaiano originando una molecola di DNA circolare Il DNA virale a doppia elica entra nelle cellule batteriche durante l’infezione come molecola lineare Cicli ripetuti di replicazione con il meccanismo del cerchio rotante Concatameri di copie di DNA virale 5’ Il DNA codifica per le proteine della testa e della coda del virus Il DNA viene inserito nelle teste del virus e tagliato a livello del sito cos Sono aggiunte le code Particella virale infettiva. I virus sono rilasciati per lisi delle cellule e possono infettare altre cellule batteriche. IMPACCHETTAMENTO del DNA VIRALE
COSMIDI Sono PLASMIDI, ma contengono, oltre alle caratteristiche essenziali di un plasmide, anche un SITO COS La presenza di questo elemento è determinante per consentire l’inserimento della molecola di DNA ricombinante nella testa del virus che, a sua volta, la inietta in una cellula batterica per infezione.
c) cosmidi Sono plasmidi che utilizzano la capacità del batteriofago lambda di impacchettare lunghi frammenti di DNA lineare all’interno del capside, di aderire alla superficie delle cellule batteriche e di iniettare il loro contenuto all’interno del batterio. Rispetto al batteriofago lambda, può contenere un inserto di DNA di dimensioni maggiori (45 kb). Il cosmide circolarizza e si comporta come un grande plasmide replicando il proprio genoma.
d) Vettori BAC Bacterial Artificial Chromosome Plasmidi di dimensioni enormi, consentono l’inserimento di frammenti di DNA umano fino a 300 Kb. Il DNA viene inserito nei batteri mediante processo di elettroporazione: uno shock elettrico provoca la formazione transitoria di pori sulla membrana batterica attraverso i quali passa il DNA.
Ospiti per i vettori • Le caratteristiche di un ospite di clonaggo sono: • crescita rapida • capacità di crescere in terreni di coltura poco costosi • non patogencità • capacità di essere trasformato • stabilità della coltura.
INTERVALLO (Lag phase) I batteri vengono diluiti nella coltura iniziale; la divisione procede lentamente in quanto le cellule si stanno adattando al terreno fresco. 5.0 4.5 4.0 3.5 3.0 2.5 2.0 1.5 1.0 0.5 0 4 - 5 ore Densità cellulare (OD600) FASE LOGARITMICA I batteri crescono esponenzialmente. 10 - 11 ore 0 5 10 15 20 Tempo (h) FASE STAZIONARIA La densità cellulare rimane costante. La coltura può anche entrare in una fase di declino in cui le cellule si lisano ed il DNA si degrada parzialmente. Curva di crescita di E. coli
3’-GGCTAAC 3’-GGCTAAC 5’ 3’ Schema di sequenziamento a terminazione di catena DNA stampo a singola elica Ibridazione con Il primer + [35S]dATP+dCTP,dGTP,dTT (dNTP) +Sequenasi ddATP, dNTP ddCTP, dNTP ddGTP, dNTP ddTTP, dNTP -CCG ddA -ddC -CC ddG -CCGA ddT -C ddC -CCGATT ddG -CCGAT ddT A C G T -CCGATT ddG -CCGAT ddT -CCGA ddT -CCG ddA -CC ddG -C ddC -ddC G T T A G C Direzione di lettura C Sequenza: 5’-CCGATTG