Download

1 / 24
270 likes | 469 Views
Innate Immune System. IMMUNE SYSTEM FUNCTION To Protect against “entry” from microorganisms / pathogens - Bacteria - Viruses - Fungi - Parasites - Toxins - Venoms Recognize Pathogens Eliminate Pathogens

E N D
IMMUNE SYSTEM FUNCTION To Protect against “entry” from microorganisms / pathogens - Bacteria - Viruses - Fungi - Parasites - Toxins - Venoms Recognize Pathogens Eliminate Pathogens Enhance cellular synthesis and repair processes in the face of damage (or metabolic stress) Two Basic “Systems” - Innate Immune System: non-specific response directed against “anything that does not belong” that also responds to any kind of cellular stress - Adaptive Immune System: highly specific response directed towards a single pathogen or a tiny component of a single pathogen
IMMUNE SYSTEM RESPONSE Innate - First “Line of Defense” - Non- specifically blocks or attacks pathogens Skin - provides a waterproof barrier to prevent organisms from entering our “system” Mucosal Epithelia - can trap pathogenic particles and prevent them from getting through epithelial cells to tissue Leukocytes Neutrophils, Eosinophils, Basophils & Mast Cells Macrophages NK-Cells Vascular endothelial cells
Blood vessels have very tight junctions between the cells that make up the vessel walls. Only when inflammatory signals are produced locally will pores open up in the vessel wall to allow neutrophils, monocytes, and other immune cells to squeeze out and enter the tissue. Lymph vessels have relatively large gaps in the vessel walls so the various immune cells can easily move from the lymph into the tissues and back again into the lymph. The cardiovascular system and lymph system provide a very convenient transportation system for Neutrophils, monocytes, NK cells, and other immune cells
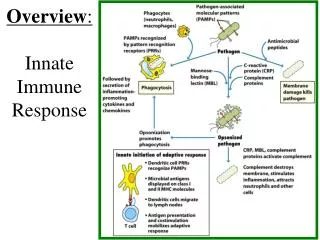
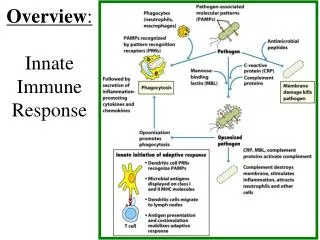
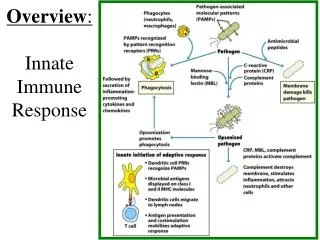
INNATE IMMUNE SYSTEM (RESPONSE) The term innate refers to the fact that these innate immune processes are not directed specifically toward any particular pathogen but rather comprise part of a very general defense which defends against any type of foreign pathogen. The innate system includes the skin and mucosal epithelial tissues. These tissues act as a “passive” physical barrier to prevent the invasion of bacteria and viruses. The outer layer of skin is comprised of several layers of dead skin cells which is simply a physical barrier. The epithelial cells lining the oral-GI tract and respiratory tracts secrete mucus which traps bacteria & viruses and thereby prevents their entry into the “body”. The mucus is either removed by coughing & spitting or by swallowing where the bacteria & viruses are destroyed by stomach acid or by digestive enzymes in the intestine. The role played by the inflammatory processes of the innate system is the active killing of those pathogens which make it through the physical barriers. Inflammation The processes of inflammation are actually considered to be a part of the innate immune system. A more modern interpretation of inflammatory responses though is that they comprise a graded stress-response system designed to enhance cellular functions to enable proper responses to cellular stresses and damage. The active killing process is performed predominantly by neutrophils, macrophages, and NK-cells. These cells serve to protect us from invading bacteria, viruses, fungi, and parasites (as well as some toxins and venoms). Neutrophils Neutrophils are a type of granulocyte (mast cells which live in our tissues, especially noticeable in the respiratory epithelium, also are granulocytes) that exists predominantly in the blood although many also are found in the lymph. Neutrophils respond to non-human cells by producing large amounts of oxygen radicals (among other toxic molecules) which kill the invading pathogens essentially by oxidizing the lipids and proteins in their cell membranes (rupturing and destroying them). Neutrophils are stimulated to do this by anything that is not smooth or which does not have a protein coat (which describes most non-human cells) as well as to any entity which has antibody proteins or compliment proteins attached to it. The antibodies and/or compliment stimulate the neutrophils to attack much more aggressively than if there were no antibodies (a very important function relating to the adaptive immune system response). In addition to killing cells quite efficiently, neutrophils also can phagocytize dead or dying cells.
INNATE IMMUNE SYSTEM (RESPONSE) Inflammation Macrophages Macrophages are cells (a form of monocyte) that “live” within our tissues as well as in the lymph. Macrophages recognize nonhuman pathogens in the same manner as neutrophils do and they are highly stimulated by antibodies and compliment as well. Monocytes usually circulate in the blood and when they are stimulated by various leukotrienes to enter the tissue they will differentiate to become macrophages. Macrophages are specialized for phagocytizing and digesting dead cells, dying cells, and foreign (non-human) cells. They also can produce oxygen radicals to kill things. As discussed below, they are one of the important types of antigen-presenting cells necessary for the adaptive immune response. Natural Killer Cells NK cells are a type of lymphocyte that is specialized at killing non-human cells (as well as many cancer cells) by lysing them. They recognize any cells which do not express the MHC as well as virus-infected cells and cells coated with antibodies. They kill cells by “injecting” proteins (porins) into their outer membranes that then cause the membranes to rupture. All of these cells (neutrophils, macrophages, NK cells) are more or less inactive (although the NK cells are more active all the time) and need to be activated by either antibodies, physical contact with pathogens, or eicosanoids and cytokines produced by proinflammatory responses. The inflammatory process can be initiated by physical damage to cells due to the effects of infectious agents, cellular damage due to mechanical or chemical disruption, or simply the overproduction of reactive oxygen species (ROS) from normal metabolic stress.
Overall Purpose of Inflammation • 1. Increased blood flow to increase “in-flow” of inflammatory and immune cells, • 2. Infiltration of the tissue with inflammatory cells to kill off infectious agents, • 3. Blood clots to stop the bleeding, • 4. Scar tissue formation to close up wounds, • 5. Increased protein synthesis to help repair the damage, • 6. Increased cell division (signals) to replace any dead cells, • 7. Initiation of an immune response resulting in the production of antibodies (and other things) . . .
Overall Purpose of Inflammation – More Details - First Line of Protection From Bad Things (microorganisms / pathogens) - Bacteria - Viruses - Fungi - Parasites - Toxins - Venoms - A variety of cells can respond to pathogens to initiate proinflammatory signaling by producing proinflammatory eicosanoids and cytokines (interleukins and chemokines) in order to initiate an inflammatory response, including dendritic cells, NK cells, neutrophils, tissue macrophages, mast cells, endothelial cells, and fibroblasts. Essentially all cells can produce proinflammatory eicosanoids in response to cellular stress and cellular damage. - Inflammatory Cells Recognize and Eliminate Pathogens: Neutrophils (circulate in the blood – chemoattraction into tissues) Macrophages (live in tissue/circulate as monocytes – chemoattraction into tissues) Kill things by eating and digesting them Kill things by releasing oxygen radicals: O2- H2O2 ·OH NK Cells (circulate in blood – peripheral tissues) Kills things using perforins; activated on by: - anything with no MHC - antibodies attached
How Does This Inflammation Thing Work? Specific receptors for pathogen associated molecular patterns (PAMPs) bind to components of pathogens and initiate signal transduction pathways that lead to the synthesis of the various proinflammatory signaling molecules. Specific receptors for damage associated molecular patterns (DAMPs) bind to components of dead (or dying) cells to activate essentially the same signal transduction pathways to initiate the synthesis of proinflammatory signaling molecules; signaling molecules that then activate the various inflammation processes: 1. P38-, ERK-MAPK, calcium, and ROS activate Phospholipase A which cuts off the fatty acid arachidonic acid from membrane phospholipids 2. Arachidonic acid is made into prostaglandins, leukotrienes, and thromboxanes: Prostaglandins: PGE-2 - causes vasodilation (among many other things) Leukotrienes: LTB-4 - attraction/activation of neutrophils Thromboxanes: TXA-2 – platelet aggregation 3. Activated macrophages, neutrophils, glial cells, dendritic cells, B-cells, fibroblasts, and endothelial cells produce a variety of interleukins and other cytokines: IL-1 → MCP-1: monocyte attraction, COX-2:PG/LT/TX IL-6 → fibrinogen production IL-8 → chemoattractant TNF → HSP-47/NFkB / AP-1 (fibrosis/protein synthesis/cell division) NFkB → ICAM
One of the major signaling process involved in initiating inflammation is the production of the eicosanoids: prostaglandins, thromboxanes, and leukotrienes from arachidonic acid. The enzyme phospholipase A2 (PLA2) is activated by either ROS, calcium, p38 MAPK and ERK-MAPK . Damage to cellular membranes allows calcium to leak through the membrane and activate PLA-2. Attacks on cellular membranes by bacterial toxins, bacterial digestive enzymes, or viral damage are common initiators of inflammation, as is physical trauma. Any kind of metabolic stress also will activate PLA-2 through calcium and ROS-mediated activation of the P38- and ERK-MAPK signal transduction pathways. Once activated, phospholipase A2 cleaves arachidonic acid from cellular membrane phospholipids. The arachidonic acid is a substrate for the enzyme cyclooxygenase-2 (COX-2 can be induced; meaning: more of it will be synthesized, by certain inflammatory cytokines); leading to enhanced synthesis and release of prostaglandins and luekotrienes. Leukotriene B4 is a potent chemotactic molecule which greatly enhances the accumulation of leukocytes within the inflammatory area while thromboxane A2 activates platelets. Prostaglandin E2 (can cause vasodilation to enhance blood flow to the infected (or damaged) area and for enhancing vasopermeability to allow neutrophils easy entry to the tissue from the blood. Thus, one end result of the production of all of these signaling molecules is the efficient recruitment and activation of neutrophils and macrophages to kill and digest any pathogens as well as dying (or dead) cells. In addition to the activation of neutrophils and macrophages, eicosanoid molecules also activate other inflammatory cells, such as glial cells, dendritic cells, B-cells, fibroblasts, as well as other endothelial cells that may be in the region. Although most of these other cells will be “ignored” here, they are still important in the inflammatory process because when they are activated to produce inflammatory signals, the result is an amplification of inflammatory signals being produced within the damaged region. Thus, they participate in a cascade of effects: damaged cells produce inflammatory signals which stimulate other cells in the local region to produce inflammatory signals, amplifying the production of inflammatory signals; resulting in the efficient recruitment of inflammatory cells into the region. (These signaling molecules have many other effects than the ones listed above... These examples simply illustrate the concept that each change in cellular function demands a signaling molecule of some sort to initiate the change in function)
Activation of PLA-2 by P38- and ERK-MAPKs, calcium, or ROS leads to removal of Arachidonic acid (or Eicosapentaenoic acid / Docosahexaenoic acid) from the phospholipid: phosphatidyl choline. These fatty acids are picked up by COX and LOX enzymes and synthesized into several prostaglandins, leukotrienes, and thromboxanes by the specific cell-types indicated, while the Lyso-PC is synthesized into Platelet Activating Factor. On binding to their appropriate receptors these lipid-derived signaling molecules initiate the responses indicated in the red boxes.
As a result of pathogen- or chemical-induced membrane damage (DAMPS), or contact with a pathogen (PAMPS) a variety of signaling pathways are activated which then stimulate the synthesis, or expression, (and extracellular release) of IL-1, IL-6, and TNF by the pathogen-activated macrophages or the damaged endothelial cells (or pathogen-activated neutrophils). These cytokines can then initiate the activation of transcription activators in many of the cells in the damaged or infected region. They can stimulate the expression (transcription and translation) of the acute-phase inflammatory response elements: cyclooxygenase-2 (COX2), IL-8, NFκβ and c-reactive protein. IL-6, TNF, and NFκβ in turn , activate other transcription activator proteins which then induce the synthesis of P-selectin, E-selectin, VCAM (vascular cell adhesion molecule), and ICAM (inter cellular adhesion molecule) by endothelial cells, especially those of the capillaries (other effects of these cytokines are discussed below). These cellular adhesion molecules are necessary for the recruitment of circulating neutrophils and monocytes into the infected or damaged area while the other interleukins activate these (and other) inflammatory cells. Once activated by the various PGs, PAMPs and/or DAMPs, neutrophils and macrophages can produce large quantities of reactive oxygen species (ROS) such as superoxide and hydrogen peroxide (other highly reactive molecules are produced but they are being ignored here) which then react with each other to produce the highly reactive (and damaging) hydroxyl radical. These ROS are responsible for killing the (pathogens). Unfortunately, they also cause oxidative damage to membranes of surrounding cells and therefore create an enhanced risk for inflammation-mediated cytotoxicity. These ROS also can cause DNA-damage in the normal cells in the inflamed region leading to enhanced risk for cancer. The activated monocytes recruited into the tissue (via adhesion molecules) mature to become macrophages (to add to the resident macrophages) to add to this inflammatory response. Another consequence resulting from the production of these signaling molecules is the formation of blood-clots and/or scar tissue to close the wound. (This could, however, be a problem if there is no wound to close.) Blood-clot formation is initiated by IL-6 which activates platelets to become sticky (clotting factors will conveniently be ignored in this course). Scar formation is initiated by several of the cytokines produced by the stimulated neutrophils and macrophages. Both IL-1 and TNF can stimulate the activation of the transcription activator AP-1 which, among other things, induces expression of TGF-β; TGF-β then stimulates tissue fibroblasts to produce HSP-47. HSP-47 mediates enhanced collagen synthesis by these fibroblasts. IL-6 stimulates both hepatocytes and epithelial cells to produce fibrinogen which, along with collagen, provides the major proteins necessary for the resulting fibrosis (scar formation).
Damage Associated Molecular Patterns (DAMPs)and Pathogen Associated Molecular Patterns (PAMPs) initiate a series of enzyme reactions that result in the activation of the p38- and ERK-MAPK signal transduction pathways. Some of the interleukins and growth factors produced as part of the inflammatory response can lead to increased rates of protein synthesis and cell division (presumably to help repair damage and to replace dead cells…)
An intact cell membrane acts as a barrier for (among other things) charged particles such as calcium. A damaged cell membrane will allow (among other things) charged particles to enter the cell or organelle.
Entry of calcium into the cytosol via normal signaling events mediated by DAG or through ROS-damaged membranes activates PKC, which then may activate PLA-2, and the ERK- and p38-MAPK pathways. At the same time calcium-activated increases in cellular metabolism subsequently increases production of ROS, which also may activate PLA-2 as well as p38-MAPK and JNK-MAPK via Trx-Ask1. ROS-caused single and double-strand breaks in DNA activates repair by ATM which also activates NFκβ and ROS-mediated PI3K activity also enhances mTORC1 via Akt to enhance overall rates of protein synthesis. The result of all these effects is the synthesis of a variety of proinflammatory molecules in response to cellular stress.
Another cellular response initiated by the inflammatory signals is stem cell division. When there is sufficient damage to kill tissue cells, they then need to be replaced. Stem cells in the tissue respond to the various inflammatory cytokines by increasing their rates of protein synthesis in order to grow and then they divide, producing daughter cells which replace the dead tissue cells. Signaling molecules called growth factors are important in producing this cell growth/cell division effect. The chemokine CXCL8 (formerly known as IL-8) stimulates platelets (which are activated by IL-6) to synthesize and release platelet-derived growth factor (PDGF) which stimulates all cells in the region to enhance rates of protein synthesis. Prolonged activation of the transcription activator AP-1 also leads to greatly enhanced rates of protein synthesis and ultimately (in stem cells) to cell division. Thus the inflammatory cytokines produced by stressed or damaged cells and then by stimulated macrophages and neutrophils are responsible for initiating all of the steps necessary for repair of wounds and protection from infection: enhanced rates of protein synthesis for cell-repair, enhanced rates of fibrosis for wound healing, enhanced rates of stem cell division to replace dead cells, and activation of neutrophils, macrophages, and dendritic cells to kill any invading pathogens. This last result of the primary inflammatory response is very important in initiating the development of an adaptive immune response. The adaptive immune response ultimately results in the production of antibodies which recognize specific antigens. Antibodies are proteins which are produced by antibody-producing B-lymphocytes. These antibody proteins specifically bind to structures that are found only on the pathogen which initiated the adaptive response in the first place. One antibody produced by one B-lymphocyte binds to one antigen while a different antibody produced by a different B-lymphocyte binds to a different antigen. One pathogen (let’s say a particular strain of bacteria such as Mycobacterium tuberculosis) may be capable of having hundreds of different antigenic sites as part of the bacterium. One antigenic site may be composed of several amino acids from a particular protein that corresponds to a shape that is definitely “not human” in origin. With several hundred different proteins making up the bacterium it is possible that there could be hundreds of short sequences of amino acids among those bacterial proteins which possibly represent different antigenic sites. For many other strains of bacteria, some of the lipids contained in the bacterial cell wall are so different from those in humans that they also might be capable of being antigens. How these antigens actually stimulate the adaptive immune response is a real cool process; a process which starts with antigen-presenting cells such as the macrophage or dendritic cell.
Regulation of Tissue Regeneration PGE2 and other growth factors activate various MAPK signal-transduction pathways in Stem Cells to initiate the activation and synthesis of a variety of proteins that are necessary for activating growth of the stem cell and the synthesis of DNA to duplicate the chromosomes in order to proceed to the mitosis/cytokinesis phase. A decline in the levels of PGE2 and of growth factors induces G0 arrest via reducing MAPK activities. Progenitor cells divide more-or-less continually in response to normal levels of stress-response signaling with the displaced daughter cells differentiating into tissue cells. In skin and intestine extensive tissue damage greatly enhances rates of cell division in stem cells and their daughter cells can differentiate directly into tissue cells.
Some of the recruited immune cells produce reactive oxygen species which may then cause damage to other cells in the region…
With an appropriate inflammatory response bacteria are killed before they can cause extensive damage and the wound is closed and tissue repaired before additional infections can occur and cause more damage.
A Brief Review: As a result of all of the inflammatory signaling molecules being produced, a variety of cellular responses occur in the affected tissues: 1. Increased blood flow to increase “in-flow” of inflammatory and immune cells, 2. Infiltration of the tissue with inflammatory cells to kill off infectious agents, 3. Blood clots to stop the bleeding, 4. Scar tissue formation to close up wounds, 5. Increased protein synthesis to help repair the damage, 6. Increased cell division (signals) to replace any dead cells, 7. Initiation of an immune response resulting in the production of antibodies (and other things) . . . So . . . Exactly why do I want to know all this?
… why is all this inflammation and cell-function stuff important? Well, we are made up of billions of cells and the proper function of our organs (each made up of a different array of specialized cells) depends on the proper functioning of the individual cells. Normal processes of metabolism in cells are the processes that ensure proper cell function. While these processes are highly regulated, they can be increased by any additional calcium entering the cell. Normal metabolic processes produce reactive oxygen specie (ROS) and therefor any additional metabolism will result in the production of small amounts of additional reactive oxygen species ROS. Normal functions of the protein synthesis enzymes and the lipid synthesis enzymes and the repair enzymes are regulated by a variety of signaling molecules as well as by ROS and calcium through their combined effects on signal transduction pathways; ensuring that any increase in damage from ROS due to any cell stress is repaired and cell function continues. When there is damage from, say, a cut to the skin, liver damage from drinking a little turpentine, or lung damage from breathing in smoke from burning plastic, or damage to respiratory-tract tissues from a bacterial or viral infection, then an inflammatory response will occur to accelerate the repair/replace processes to ensure that the damage is repaired, dead cells are replaced, and tissue (or organ) function is returned to normal. Because normal signal transduction processes are regulated in part by both calcium and ROS; this means that chronic elevation in both because of chronic stress to cells will result on chronic over-activation of these S-T pathways… … resulting in chronic proinflammatory signaling
The chronic proinflammatory signaling can then lead to the invasion of inflammatory cells into the tissue that then cause lots of ROS-mediated damage and much more proinflammatory signaling. The damage can of course, lead to cell death. If only a few cells die then nobody cares… but if a whole bunch of cells in a single organ die at once, we just might be in trouble: if a bunch of heart or brain cells die, we may go along with them. If a bunch of liver cells die, we may suffer liver failure… if it is “mild” the inflammatory response may save our liver. The rest of our cells in our other organs however, suffer until the liver is completely repaired because they depend on the liver to produce many different molecules that are necessary for their proper function and to handle the wastes produced by all of our cells. If the liver damage is too severe to repair quickly, then other organs can fail because they don’t get what they need and again, we will die along with them. In essence, cells in each of the different organs are all inter-dependent on the function of all the other cells in other organs. Obviously without the inflammation response to start the repair & replace process you would die at the first scratch… on the other hand, anything that inappropriately enhances this response, or sustains it for long periods of time such as: overexposure to toxic chemicals (cigarette smoke, toluene, carbon tetrachloride, ethanol, chlorine…) or to a lack of specific nutrients and beneficial phytochemicals… inevitably leads to continual local production of growth signals and ROS from the inflammatory cells and eventually a slow & painful death from organ failure, cardiovascular disease, cancer, or some other degenerative disease… Ultimately, our health depends on optimal function of our cells! And this function can be impaired by aberrant inflammation.
The only way to stop proinflammatory signaling is to produce the proresolution signaling molecules that then stop the production of the proinflammatory signaling molecules. Proinflammatory signaling leads to an induction of COX-2 and mPGES; increasing PGE-2 synthesis and subsequently an enhanced expression of 12-LOX and 15-LOX. Increased LOX in conjunction with COX-2 greatly enhances the synthesis of lipoxins, resolvins, and PGJ-2. Lipoxins and resolvins suppress proinflammatory signaling via inhibiting the activation of MAPKs and NFκβ through enhanced expression of SOCS in all inflammatory cells while PGJ-2 activates apoptosis in neutrophils as well as suppresses MAPKs and NFκβ. At the same time, macrophages phagocytize apoptotic neutrophils (without production of ROS) and are stimulated to egress from the tissue, effectively ending an inflammatory response.