Download
1 / 53
540 likes | 747 Views
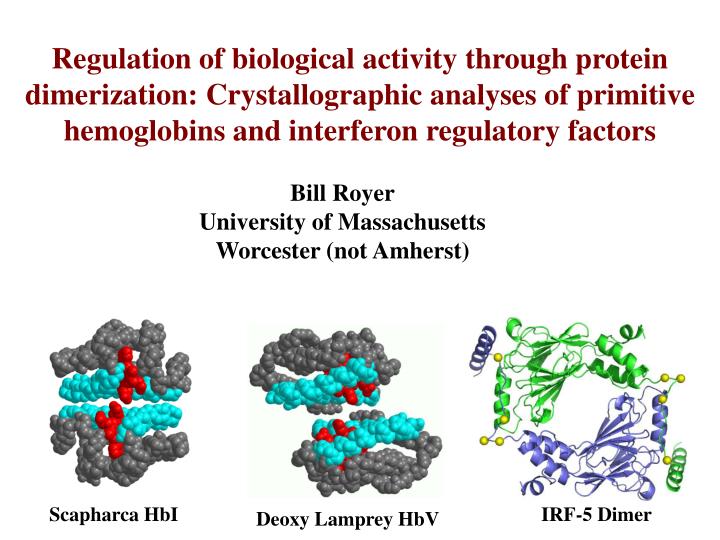
Regulation of biological activity through protein dimerization: Crystallographic analyses of primitive hemoglobins and interferon regulatory factors. Bill Royer University of Massachusetts Worcester (not Amherst). Scapharca HbI. IRF-5 Dimer. Deoxy Lamprey HbV. Molluscan hemoglobin subunit.

E N D
Regulation of biological activity through protein dimerization: Crystallographic analyses of primitive hemoglobins and interferon regulatory factors Bill Royer University of Massachusetts Worcester (not Amherst) Scapharca HbI IRF-5 Dimer Deoxy Lamprey HbV
Molluscan hemoglobin subunit Sperm whale myoglobin (Structure by John Kendrew, illustration by Irving Geis)
Lumbricus terrestris Erythrocruorin (180 subunits) Riftia pachyptila C1 Hb (24 subunits) Annelida Homo sapiens HbA (tetramer) Echiura Urechis caupo Hb (tetramer) Caudina arenicola HbD (dimer) Chordata Echinodermata Mollusca Petromyzon marinus HbV (deoxy dimer) Protostomes Deuterostomes S. inaequivalvis HbI (dimer) Scapharca inaequivalvis HbII (tetramer)
Deoxy HbI – 1.6 Å resolution HbI-CO – 1.4 Å resolution
Cooperative ligand binding in HbI relies on only small subunit rotations Unlike binding of oxygen to human hemoglobin crystals, binding of oxygen to HbI crystals is fully cooperative (Mozzarelli et al, 1996). Despite rather localized structural transitions, the R-state is estimated to bind oxygen 300 times more tightly than the T-state.
p50 (torr) n WT 10.0 1.5 Mutation of Phe 97 leads to increased oxygen affinity and sharply diminished cooperativity F97L 1.0 1.2 F97A 0.3 1.2 F97Y 0.08 1.2 Ligation of HbI results in extrusion of Phe 97 from the proximal pocket Distal His Proximal His Phe 97 Distal His ligand Proximal His Phe 97
Deoxy T72V p50=0.2 Torr, n=1.7 The isosteric mutation Thr E10 to Val reveals the importance of the observed water cluster for stabilization of the low affinity state Deoxy HbI p50=10 Torr, n=1.5
Raising the osmotic pressure increases the oxygen affinity of HbI, supporting a key role of water molecules in stabilizing the low affinity state of HbI
Restricting heme movement abolishes allosteric transition Mutation of I114F results in low affinity and cooperativity (n=1.05, p50 = 21 Torr). 114F I114F 114F 114I HbI 114I
Mutation of distal His to Gln abolishes heme movement and allosteric transition E7Gln E7Gln H69Q E7His E7His HbI
Key structural transitions with functional ramifications Heme movement What is the cascade of structural events? F4 Phe flipping Are these transitions concerted or sequential? Interface water rearrangment Do structural kinetic intermediates facilitate R to T transition?
Time-resolved crystallography to obtain snapshots along the trajectory between high-affinity and low affinity states BIOCARS 14-IDB, APS Heat load shutter ms shutter Single bunch ~150ps X-rays (Undulator) CCD 2 s chopper e- Cycle time – 3.683 ms Dye laser Superbunch ~500ns Experimental Setup
E E F F F4 (Red: -2.5s Blue: 2.5s) CD CO* F CD1 E7 CO CD3 5ns Fe F8 F F4 F7 (Red: -2.5s Blue: 2.5s) F4 CD E7 F CD1 60ms CD3 Fe F F8 F4 F7 Subunit A Fo(light)-Fo(dark) - (Red: -3s, Blue: 3s)
Results of difference Fourier refinement, Fo(light) –Fo(dark) coefficients
Integrated difference [Fo(light)-Fo(dark)] electron density values
Change in iron position, based on difference refinement Distance along the parallel and perpendicular components of the heme plane
Key structural transitions with functional ramifications What is the cascade of structural events? Intermediate is formed rapidly (<5ns) upon ligand release, relaxing to T-like structure in microsecond time domain Heme movement F4 Phe flipping Are these transitions concerted or sequential? Key allosteric changes appear to be tightly coupled. Interface water rearrangment Do structural intermediates facilitate R to T transition? Rapid disordering of water molecules H-bonded to propionates appears to lay the foundation for subsequent heme movement.
Deoxygenated lamprey hemoglobin oligomerizes in a proton dependent fashion, conferring a strong Bohr effect Scapharca HbI Deoxy Lamprey HbV
E’-helix N79 W72’ H73’ E75 E75’ H73 W72 N79’ E-helix The primary contacts in the deoxy lamprey HbV dimer involve the E-helices
Dimerization of Lamprey Hb sterically restricts ligand binding Proximal Histidine Distal Histidine
Lamprey hemoglobin has a very pronounced Bohr effect From: Antonini et al. (1964) Arch. Biochem. Biophys. 105, 404-408, courtesy of Austen Riggs
H73’ R71’ E31 E75’ E75 E31’ R71 H73 The strong Bohr effect in lamprey HbV can be accounted for by a cluster of glutamate residues in the interface Mutations E75Q, Y30H and H73Q support the functional importance of this dimeric structure (Y.Qiu et al. A.F. Riggs (2000) JBC 275, 13517-13528)
O-O 2O 2 - - o o o o o o o o o o o o o o o o + 2H - - - - H O-O H Linkage of proton and oxygen binding in Lamprey Hemoglobin
IFN-/a virus IFNAR Tyk2 Jak1 Stat2 IRF3 Stat1 IRF3 IRF3 IRF3 IRF3 IRF3 IRF-9 (ISGF3) P P IRF3 P IRF3 IFN- gene IRF3 other cytokines, anti-viral genes IFN- genes P IRF3 IFN- IFN- Role of Interferon Regulatory Factors (IRFs) in Immediate and Delayed Anti-Viral Responses
IBs 26S proteasome IRAK4 IRAK1 TRAF6 TAK1 3 2 1 Ub Ub Ub IBs Ub TABs MAPKs IKK IKK NF-B P P P P IKK Innate immunity is triggered by the recognition of “pathogen-associated molecular patterns” such as viral nucleic acids by Toll-like receptors (TLR) or cytoplasmic receptors. Cytoplasm virus ssRNA dsDNA TLR7, TLR8 TLR9 Endosome MyD88 MyD88 IKK TBK1 TRAF6 ? IRF5 IRF7 IRF7 IRF5 IRF7 IRF5 ATF2/c-Jun NF-B NF-B Nucleus Inflammatory cytokines IFNs IFN
P P P P P P P DD IRFs are activated by phosphorylation in the C-terminal domain C N Cytoplasm Nucleus CBP or p300
IRF-3 is constitutively expressed in all cell types and acts as a molecular sentry for viral infection. Ser/Thr PO4 sites RVGGASSLENTVDLHISNSHPLSLTS 380 110 1 200 427 380 DBD IAD IRF-3 transactivation domain construct AUD NLS NES 427 173 IAD B.Y. Qin, et al. K. Lin (2003) Nat. Struct. Biol.10, 913 -921 K. Takahashi, et al. F. Inagaki (2003) Nat. Struct. Biol. 10, 922-927 Domain structure of human IRF-3
N C IRF-3 (residues 173-427) Structure of IRF-3 transactivation domain in complex with CBP supports the hypothesis that the autoinhibitory region masks CBP binding site CBP (2067-2112) N IRF-3 (residues 173-394) B.Y. Qin, et al. K. Lin (2005) Structure13, 1269-1277
RVGGASSLENTVDLHISNSHPLSLTS 222 467 IAD Domain structure of Human IRF-3 and IRF-5 405 380 IRF-3 110 1 200 427 380 DBD IAD AUD NLS NES 427 173 IRF-3 transactivation domain construct IAD * 421 455 SGELSWSADSIRLQISNPDIKDRMV IRF-5 (variant 4) 1 140 233 421 467 DBD IAD NLS NLS NES IRF-5 transactivation domain construct
IRF-5 IRF-5 S430D + CBP 450 µM IRF-5 S430D 600 600 800 IRF-5 + CBP IRF-5 WT mAU(280 nm) 400 400 600 250 µM CBP 200 200 mAU (280 nm) 400 CBP mAU (280 nm) 100 µM 0 0 200 13.0 14.0 15.0 16.0 17.0 18.0 19.0 12.0 50 µM 12.0 13.0 14.0 15.0 16.0 17.0 18.0 19.0 Volume (ml) Volume (ml) 0 13.0 14.0 15.0 16.0 17.0 Volume (ml) 450 µM IRF-5 S430D 800 600 250 µM mAU (280 nm) 400 100 µM 200 50 µM 0 13.0 14.0 15.0 16.0 17.0 Volume (ml) Size exclusion chromatography to investigate oligomerization of IRF-5 (222-467) and IRF-5 S430D Monomer Dimer
C Helix 5 N Helix 5 N Helix 3 Helix 1 Helix 4 C Helix 2 C N IRF-5 dimeric subunit IRF-3 complex with CBP IRF-3 autoinhibited monomer
N Helix 5 C C Helix 5 N IRF-5 (222-467) S430D Dimer
Helix 4 S425 S427’ S430’(D) L403 I431’ Y303 Helix 2 L433’ L307 I435’ R328 V310 D312 S436’ F279 K449’ D442’ V445’ Helix 5 Key interface residues in the IRF-5 dimer S436’ R353 R328 D442’ R353 Helix 5
Mutation of interface residues disrupt dimer formation of IRF-5 (222-467) in solution Monomer Dimer 600 IRF5-S430D & CBP 500 IRF5-S430D/V310D & CBP IRF5-S430D/R328E & CBP 400 IRF5-S436D/R328E & CBP IRF5-S430D/R353D & CBP mAU (280 nm) 300 IRF5-S430D/D442R & CBP 200 CBP 100 0 13 14 15 16 17 18 19 20 Volume (ml)
Disruption of dimerization by mutation of interface residues inhibits full length IRF-5 activation HEK293 Cells IFNb lucerferase (Fold Induction)
Helix 5 S425 S427’ S430’(D) (homologous IRF3 residue number for absolutely conserved residues) (L362) L403 I431’ Y303 Helix 2 L433’ L307 I435’ (R285) R328 V310 D312 S436’ S436’ R353 F279 R328 K449’ D442’ D442’ V445’ R353 Helix 5 Helix 5
Mutation of IRF-3 residues homologous to IRF-5 dimeric interface residues disrupts formation of the IRF-3 (173-427) dimer in solution Monomer Dimer 400 IRF3-S386D/396D & CBP IRF3-S386D/S396D/R285E & CBP IRF3-S386D/S396D/L362D & CBP 300 mAU (280 nm) CBP 200 100 0 12.0 13.0 14.0 15.0 16.0 17.0 18.0 19.0 20.0 Volume (ml)
Disruption of IRF-3 dimerization inhibits its activation HEK293 Cells Mock NDV IFNb lucerferase (Fold Induction) IFNb lucerferase (Fold Induction) Published IRF-3 mutants reinterpreted in light of our structure also support the crystallographically observed IRF-5 dimer as representing the active state of IRF-3
IRF activation, dimerization and CBP binding Phosphorylation C-term (Morphing CNS script from the Yale Morph Server, http://molmovdb.org)
IRF activation, dimerization and CBP binding Phosphorylation (Morphing CNS script from the Yale Morph Server, http://molmovdb.org)
DBD DBD Helix 5 Helix 5 Helix 5 CBP binding site CBP binding site P P P DBD P P P P P P CBP binding site CBP binding site Helix 5 Helix 5 Helix 5 DBD DBD DBD CBP DBD Helix 5 CBP binding site DBD P P DBD P P CBP binding site Helix 5 Cytoplasm Nucleus
IRF project Weijun Chen Suvana Lam Hema Srinath Brendan Hilbert Celia Schiffer Kate Fitzgerald Zhaozhao Jiang Hemoglobin projects James Knapp Holly Heaslet Animesh Pardanani Michele Bonham Candace Summerford Michael Omartian Jeff Nichols Quentin Gibson BioCARS – Univ. of Chicago Time-resolved Crystallography Vukica Srajer Reinhard Pahl $ - NIH Kai Lin
IRF-5 • Autoinhibition of IRF-5 is less tight than that for the ubiquitously expressed IRF-3. • IRF-5 is activated by: • infection by some viruses • type I interferon • tumor suppressor p53 • IRF-5 activates • type I interferon • proinflammatory cytokines, including TNF-a, IL-12 and IL-6 • tumor suppressors • Human mutations of IRF-5 have been implicated in • systemic lupus erythematosis • multiple sclerosis • Sjogrens syndrome • Inflamatory bowel disease • IRF-5 k.o. mice show • susceptibility to viral infection • resistance to endotoxin shock • susceptibility to tumors
Interactions of CBP (2067-2112) with IRF-5 (222-467) and phosphomimetic mutants based on ITC data Complex Kd Change in affinity CBP – IRF-5 1.64mM 1.0 fold CBP – IRF-5 (S427D) 0.96mM 1.7 fold CBP – IRF-5 (S425D) 0.71mM 2.3 fold CBP – IRF-5 (S436D) 0.67mM 2.4 fold CBP – IRF-5 (S430D) 0.56mM 2.9 fold