Download
1 / 1
10 likes | 126 Views
Clusters and Superclusters of Phased Small RNAs in the Developing Inflorescence of Rice Lewis Bowman 1 , Anna Kasprzewska 1 , Vicki Vance 1 , John Fernandes 2 , Guo-Ling Nan 2 , Virginia Walbot 2 , Cameron Johnson 3 , Kristin Tennessen 3 , Venkatesan Sundaresan 3

E N D
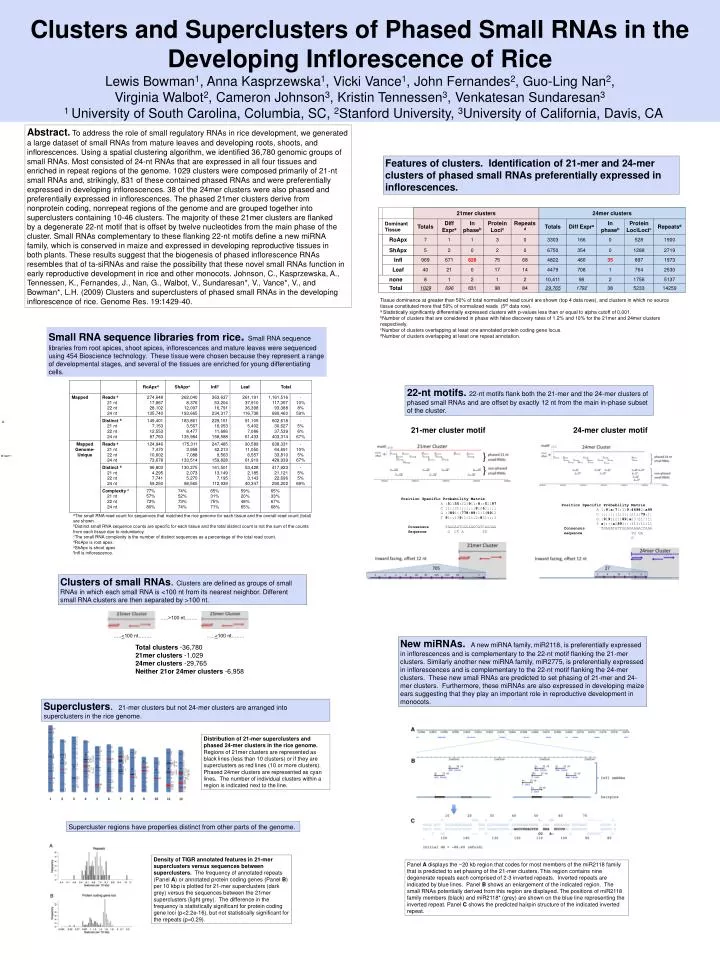
Clusters and Superclusters of Phased Small RNAs in the Developing Inflorescence of Rice Lewis Bowman1, Anna Kasprzewska1, Vicki Vance1, John Fernandes2, Guo-Ling Nan2, Virginia Walbot2, Cameron Johnson3, Kristin Tennessen3, Venkatesan Sundaresan3 1 University of South Carolina, Columbia, SC, 2Stanford University, 3University of California, Davis, CA …..>100 nt…….. …..<100 nt…….. …..<100 nt…….. Total clusters -36,780 21mer clusters -1,029 24mer clusters -29,765 Neither 21or 24mer clusters -6,958 Abstract.To address the role of small regulatory RNAs in rice development, we generated a large dataset of small RNAs from mature leaves and developing roots, shoots, and inflorescences. Using a spatial clustering algorithm, we identified 36,780 genomic groups of small RNAs. Most consisted of 24-nt RNAs that are expressed in all four tissues and enriched in repeat regions of the genome. 1029 clusters were composed primarily of 21-nt small RNAs and, strikingly, 831 of these contained phased RNAs and were preferentially expressed in developing inflorescences. 38 of the 24mer clusters were also phased and preferentially expressed in inflorescences. The phased 21mer clusters derive from nonprotein coding, nonrepeat regions of the genome and are grouped together into superclusters containing 10-46 clusters. The majority of these 21mer clusters are flanked by a degenerate 22-nt motif that is offset by twelve nucleotides from the main phase of the cluster. Small RNAs complementary to these flanking 22-nt motifs define a new miRNA family, which is conserved in maize and expressed in developing reproductive tissues in both plants. These results suggest that the biogenesis of phased inflorescence RNAs resembles that of ta-siRNAs and raise the possibility that these novel small RNAs function in early reproductive development in rice and other monocots. Johnson, C., Kasprzewska, A., Tennessen, K., Fernandes, J., Nan, G., Walbot, V., Sundaresan*, V., Vance*, V., and Bowman*, L.H. (2009) Clusters and superclusters of phased small RNAs in the developing inflorescence of rice. Genome Res. 19:1429-40. Features of clusters. Identification of 21-mer and 24-mer clusters of phased small RNAs preferentially expressed in inflorescences. Tissue dominance at greater than 50% of total normalized read count are shown (top 4 data rows), and clusters in which no source tissue constituted more that 50% of normalized reads (5th data row). a Statistically significantly differentially expressed clusters with p-values less than or equal to alpha cutoff of 0.001. bNumber of clusters that are considered in phase with false discovery rates of 1.2% and 10% for the 21mer and 24mer clusters respectively. cNumber of clusters overlapping at least one annotated protein coding gene locus. dNumber of clusters overlapping at least one repeat annotation. Small RNA sequence libraries from rice. Small RNA sequence libraries from root apices, shoot apices, inflorescences and mature leaves were sequenced using 454 Bioscience technology. These tissue were chosen because they represent a range of developmental stages, and several of the tissues are enriched for young differentiating cells. 22-nt motifs.22-nt motifs flank both the 21-mer and the 24-mer clusters of phased small RNAs and are offset by exactly 12 nt from the main in-phase subset of the cluster. A 21-mer cluster motif 24-mer cluster motif Position Specific Probability Matrix A :51155:221911:8::51197 C :1::31:::::::8:26::::: G :3891::778:89:1:149912 T 91::139:1:11:2:821:::1 Consensus TAGGAATGGGAGGCATCAGGAA Sequence G CT A TG Position Specific Probability Matrix A 1:91a:71:119:46982:a99 C :::::::1:1:::211:79:1: G :91911211891a23:21::11 T a1:::a2891:::311:11:11 Consensus TGAGATATTGGAGAAAACCAAA sequence TG GA G aThe small RNA read count for sequences that matched the rice genome for each tissue and the overall read count (total) are shown. bDistinct small RNA sequence counts are specific for each tissue and the total distinct count is not the sum of the counts from each tissue due to redundancy. cThe small RNA complexity is the number of distinct sequences as a percentage of the total read count. dRoApx is root apex. eShApx is shoot apex. fInfl is inflorescence. Clusters of small RNAs. Clusters are defined as groups of small RNAs in which each small RNA is <100 nt from its nearest neighbor. Different small RNA clusters are then separated by >100 nt. New miRNAs. A new miRNA family, miR2118, is preferentially expressed in inflorescences and is complementary to the 22-nt motif flanking the 21-mer clusters. Similarly another new miRNA family, miR2775, is preferentially expressed in inflorescences and is complementary to the 22-nt motif flanking the 24-mer clusters. These new small RNAs are predicted to set phasing of 21-mer and 24-mer clusters. Furthermore, these miRNAs are also expressed in developing maize ears suggesting that they play an important role in reproductive development in monocots. Superclusters. 21-mer clusters but not 24-mer clusters are arranged into superclusters in the rice genome. Distribution of 21-mer superclusters and phased 24-mer clusters in the rice genome. Regions of 21mer clusters are represented as black lines (less than 10 clusters) or if they are superclusters as red lines (10 or more clusters). Phased 24mer clusters are represented as cyan lines. The number of individual clusters within a region is indicated next to the line. Supercluster regions have properties distinct from other parts of the genome. Density of TIGR annotated features in 21-mer superclusters versus sequences between superclusters. The frequency of annotated repeats (Panel A) or annotated protein coding genes (Panel B) per 10 kbp is plotted for 21-mer superclusters (dark grey) versus the sequences between the 21mer superclusters (light grey). The difference in the frequency is statistically significant for protein coding gene loci (p<2.2e-16), but not statistically significant for the repeats (p=0.29). Panel A displays the ~20 kb region that codes for most members of the miR2118 family that is predicted to set phasing of the 21-mer clusters. This region contains nine degenerate repeats each comprised of 2-3 inverted repeats. Inverted repeats are indicated by blue lines. Panel B shows an enlargement of the indicated region. The small RNAs potentially derived from this region are displayed. The positions of miR2118 family members (black) and miR2118* (grey) are shown on the blue line representing the inverted repeat. Panel C shows the predicted hairpin structure of the indicated inverted repeat.