Download

1 / 63
640 likes | 908 Views
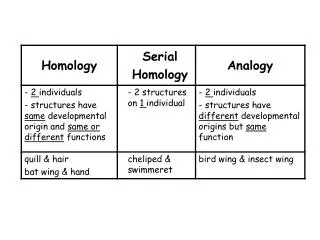
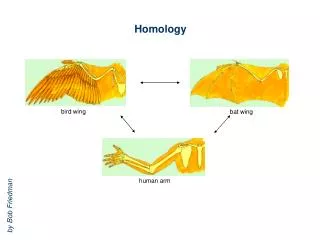
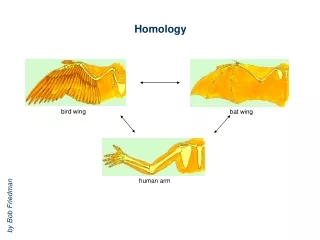
Developmental homology and dissociation. Homologous genes need not function in the development of homologous structures (HOX genes, Notch signaling) Expression of a homologous gene does not imply that developmental pathways are also homologous ( engrailed and metamerism)

E N D
Developmental homology and dissociation Homologous genes need not function in the development of homologous structures (HOX genes, Notch signaling) Expression of a homologous gene does not imply that developmental pathways are also homologous (engrailed and metamerism) Homologous developmental pathways may control the development of non-homologous structures (Dll in appendages, Pax6 in the eyes) Homologous structures need not be specified by homologous genes (insect segmentation)
Segment polarity genes make up the bottom level of the regulatory hierarchy Segment polarity genes establish boundaries between segments and control patterning within each segment
Expression of segment polarity genes is conserved in beetles… en wg Tribolium
Grasshoppers en Schistocerca
Myriapods en/wg Lithobius
Crustaceans Artemia wg
Spiders en Cupiennius salei wg The functions of en and wg in subdividing the embryo into segments appear to be conserved
eve pair-rule function is conserved in beetles Chromophore-assisted laser inactivation
ftz deletion does not affect segmentation in Tribolium Antennae
Pair-rule gene expression in Schistocerca Drosophila Tribolium eve ftz pby
even-skipped expression in Lithobius eve/ en eve expression does not show two-segment periodicity
Expression of pair-rule genes in Chelicerates primary pair-rule genes in Cupiennius paired - secondary pair-rule in Tetranychus
Evolution of Arthropod segmentation - Some parts of the segmentation pathway are conserved. - There is some turnover of genes within the overall pathway - Pair-rule patterning may be a higher insect innovation
Segmentation in long germ band insects • Simultaneous generation of segments • Segmentation independent of growth • - Occurs in syncytial environment
Cellularization before blastoderm formation in grasshoppers Rhodamine dextran injection
Segmentation in short germ band insects (Tribolium) • Sequential generation of segments • Segmentation coupled to growth • - Occurs in a cellular environment The global patterning mechanisms cannot operate in the same way as in Drosophila
Maternal gradient of bicoid establishes Anterior-Posterior axis in the Drosophila embryo
The roles of maternal gradients in Drosophila Gradients form from maternally deposited transcripts by diffusion or transport in a cell-free environment
bicoid function is conserved in Cyclorhapha Maternal gradients in Drosophila and Megaselia RNA interference Inhibition of Bcd protein synthesis in Megaselia results in posterior duplication (an embryo with two butts…)
hunchback is a conserved component of the Anterior determination system Megaselia RNA interference Schistocerca
Tribolium hunchback is correctly regulated in Drosophila Tribolium hb Tribolium hb transgene in Drosophila Some maternal system must therefore exist in beetles But how does it work without bicoid?
hunchback can substitute for bicoid Making anterior hunchback stripe in the absence of bicoid
In Tribolium, anterior patterning is controlled by orthodenticle and hunchback otd hb otd; hb otd is deposited maternally Removal of otd and hb eliminates anterior structures
A new mechanism for a new mode of development Drosophila Tribolium bcd hb otd Maternal hb otd hb Zygotic Anterior structures Anterior structures A maternal protein gradient can only work in a syncytium bcd has taken over the ancestral functions of otd and hb ?
A parasitic wasp, Copidosoma floridanum Polyembryonic development
Polyembryonic development Primary morula Polymorula Secondary morulae
Polyembryonic development evolved independently as an adaptation to parasitism
Segmentation without maternal gradients engrailed expression in Aphidius ervi
Segmentation without pair-rule genes? eve eve Bracon Aphidius en
How do developmental pathways diverge from a common ancestral state? • By recruitment and loss of component genes • By re-deployment of old genes in new patterns • By changing regulatory interactions between genes
Somatic sex determination pathway in Drosophila Sex determination is cell-autonomous (X:A ratio or dsx expression)
Somatic sex in Drosophila is controlled by a splicing cascade Establishment (X:A ratio) Maintenance (autoregulation)
Regulation of downstream target genes by doublesex Yolk protein expression Genotype/sex
Sex determination mechanisms in insects Y-chromosomal genes (Tipulidae, Tephritidae) Autosomal genes (Culex, Anopheles) Mobile genes (Megaselia, Musca) X:Autosome ratio (Drosophila) Genotype of the mother (Chrysomia, Sciara) Haploid/ diploid (Hymenopterans) Environmental factors (Pseudacteon)
Sex determination in the medfly Ceratitis capitata (Tephritidae) Sxl Sex is controlled by a male-determining factor on the Y tra
transformer controls sexual differentiation in Ceratitis Female Male Intersexes produced by tra RNAi
Sex determination in Megaselia Megaselia lacks differentiated sex chromosomes The Maleness factor is mobile and can be located on different chromosomes This can create new Y chromosomes from former autosomes
Sex determination systems in Musca domestica In male-heterogametic strains, sex is determined by a single masculinizing factor (M), which can be located either on a Y chromosome, or on 4 different autosomes Some female-heterogametic strains are homozygous for M, and sex is determined by a dominant feminizing factor F Other female-heterogametic strains lack M, and sex is determined by a recessive masculinizing factor Fman In arrhenogenic strains, sex of the offspring depends on the genotype of the mother
Musca doublesex expression is sexually dimorphic Somatic sex correlates with sexually dimorphic doublesex splicing, irrespective of the upstream sex determination mechanism