Download
1 / 26
280 likes | 516 Views
1987. Daniel Amit 1938-2007. (Ising model) ferromagnetic. (e.g., S.K. model) + spin-glass state. (Hopfield model) + memory states. Systems of spin-like elements may dynamically relax governed by the Hamiltonian towards increasingly complex “discrete” attractor states. A. B. C.

E N D
1987 Daniel Amit 1938-2007
(Ising model) ferromagnetic (e.g., S.K. model) + spin-glass state (Hopfield model) + memory states Systems of spin-like elements may dynamically relax governed by the Hamiltonian towards increasingly complex “discrete” attractor states A B C D
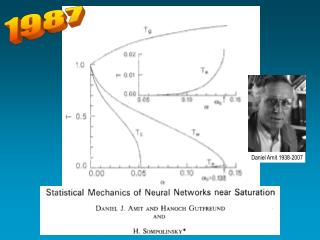
Daniel Amit, Hanoch Gutfreund and Haim Sompolinsky showed how to extend the mathematical analysis of spin-glasses into a new “statistical physics” of Hopfield attractor networks No. Almost.
They derived the free-energy and the saddle-point equations that describe the equilibrium reached by relaxational dynamics DS (disordered state) SG (spin glass) yielding finally a phase-diagram +RS (retrieval memory state) storage load = p / N
AT, J. Phys. A: Math. Gen. 24 (1991) 2645-2654. are spin-glass effects really relevant to understanding realistic auto-associative networks? binary threshold-linear: Threshold-linear neural network (Hopfield) Threshold-linear spin glass (SK) No. Almost.
Camillo Golgi 1843-1926 Teoria reticolare (il sincizio) Santiago Ramón y Cajal 1852-1934 Teoria cellulare (i neuroni) Premio Nobel 1906 La reazione nera 1873
ArealizationandMemory in the Cortex monkey Main perspectives: a) Content-based b) Hierarchical c) Statistical/modular
The statistical/modular perspective The Braitenberg model N pyramidal cells √N compartments √N cells each A pical synapses B asal synapses
A simple semantic network (O’Kane & Treves, 1992) updated to remove the ‘memory glass’ problem (Fulvi Mari & Treves, 1998) Reduced to a Potts model (Kropff & Treves, 2005) Cortical modules Structured long-range connectivity Potts units with dilute connectivity ..but all cortical modules share the same organization… Local attractor states (=S) “0” state included S+1 Potts states Global activity patterns Sparse global patterns Sparse Potts patterns pc S ?!?! pc CS 2 !!
The numbers of expansion recodingç Each MF terminates in several hundreds rosettes Each rosette has the dendrites of 28 GCs Each GC receives from 4 rosettes (MFs) There are 450 times more GCs than MFs In humans, there are 3x1010 GCs, each making about 300 PF synapse, for a total 1013 storage locations on some 5x107 Purkinje cells.
Nature411, 189 - 193 (2001); doi:10.1038/35075564 < p> Nature 411, 189 - 193 (2001) Scalable architecture in mammalian brains DAMON A. CLARK*†, PARTHA P. MITRA‡ & SAMUEL S.-H. WANG* * Department of Molecular Biology and† Department of Physics, Princeton University, Princeton, New Jersey 08544, USA‡ Bell Laboratories, Lucent Technologies, 600 Mountain Avenue, Murray Hill, New Jersey 07974, USA Correspondence and requests for materials should be addressed to S.S.-H.W. (e-mail: samwang@molbio.princeton.edu). Comparison of mammalian brain parts has often focused on differences in absolute size, revealing only a general tendency for all parts to grow together. Attempts to find size-independent effects using body weight as a reference variable obscure size relationships owing to independent variation of body size and give phylogenies of questionable significance. Here we use the brain itself as a size reference to define the cerebrotype, a species-by-species measure of brain composition. With this measure, across many mammalian taxa the cerebellum occupies a constant fraction of the total brain volume (0.13 0.02), arguing against the hypothesis that the cerebellum acts as a computational engine principally serving the neocortex. Mammalian taxa can be well separated by cerebrotype, thus allowing the use of quantitative neuroanatomical data to test evolutionary relationships. Primate cerebrotypes have progressively shifted and neocortical volume fractions have become successively larger in lemurs and lorises, New World monkeys, Old World monkeys, and hominoids, lending support to the idea that primate brain architecture has been driven by directed selection pressure. At the same time, absolute brain size can vary over 100-fold within a taxon, while maintaining a relatively uniform cerebrotype. Brains therefore constitute a scalable architecture.
paleocortex (olfactory) archicortex (hippocampus) neocortex (the rest)
Brain Behav Evol. 1997;49(4):179-213. The telencephalon of tetrapods in evolution.Striedter GF.Department of Psychobiology, University of California, Irvine 92697-4550, USA.Numerous scientists have sought a homologue of mammalian isocortex in sauropsids (reptiles and birds) and a homologue of sauropsid dorsal ventricular ridge in mammals. Although some of the proposed theories were enormously influential, alternative theories continued to coexist, primarily because the striking differences in pallial organization between adult mammals, sauropsids, and amphibians enabled different authors to enlist different subsets of similarity data in support of different hypotheses of putative homology. A phylogenetic analysis based on parsimony cannot discriminate between such alternative hypotheses of putative homology, because sauropsids and mammals are sister groups. One solution to this dilemma is to include embryological patterns of telencephalic organization in the comparative analysis. Because early developmental stages in different taxa tend to resemble each other more than the adults do, the embryological data may reveal intermediate patterns of organization that provide unambiguous support for a single hypothesis of putative homology. The validity of this putative homology may then be supported by means of a phylogenetic analysis based on parsimony. A comparative analysis of pallial organization that includes embryological data suggests the following set of homologies. The lateral cortex in reptiles is homologous to the piriform cortex in birds and mammals. The anterior dorsal ventricular ridge in reptiles is probably homologous to the neostriatum and ventral hyperstriatum in birds and to the endopiriform nucleus in mammals. The posterior dorsal ventricular ridge in reptiles is most likely homologous to the archistriatum in birds and to the pallial amygdala in mammals. The pallial thickening in reptiles is probably homologous to the dorsal and intercalated portions of the hyperstriatum in birds and to the claustrum proper in mammals. Finally, the dorsal cortex in reptiles is probably homologous to the accessory hyperstriatum and parahippocampal area in birds and to the isocortex in mammals. These hypotheses of homology imply relatively minor evolutionary changes in development but major changes in neuronal connections. Most significantly, they imply the independent elaboration of thalamic sensory projections to derivatives of the lateral and dorsal pallia in sauropsids and mammals, respectively. They also imply the independent evolution of lamination in the pallium of birds and mammals.
Biochem Cell Biol. 1997;75(6):651-67. • The brain in evolution and involution.Parent A.Laboratoire de neurobiologie, Universite Laval Robert-Giffard, Beauport, QC, Canada.This paper provides an overview of the phylogenetic evolution and structural organization of the basal ganglia. These large subcortical structures that form the core of the cerebral hemispheres directly participate in the control of psychomotor behavior. Neuroanatomical methods combined with transmitter localization procedures were used to study the chemical organization of the forebrain in each major group of vertebrates. The various components of the basal ganglia appear well developed in amniote vertebrates, but remain rudimentary in anamniote vertebrates. For example, a typical substantia nigra composed of numerous dopaminergic neurons that project to the striatum already exists in the brain of reptiles. Other studies in mammals show that glutamatergic cortical inputs establish distinct functional territories within the basal ganglia, and that neurons in each of these territories act upon other brain neuronal systems principally via a GABAergic disinhibitory output mechanism. The functional status of the various basal ganglia chemospecific systems was examined in animal models of neurodegenerative diseases, as well as in postmortem material from Parkinson's and Huntington's disease patients. The neurodegenerative processes at play in such conditions specifically target the most phylogenetically ancient components of the brain, including the substantia nigra and the striatum, and the marked involution of these brain structures is accompanied by severe motor and cognitive deficits. Studies of neural mechanisms involved in these akinetic and hyperkinetic disorders have led to a complete reevaluation of the current model of the functional organization of the basal ganglia in both health and disease.
Neocortex (+)n+ Lamination, Arealization Who is cutting-edge, in cerebellar technology? Hippocampus (+)n+ DG input sparsifier CA1 feed-forward Cerebellum ++- Expansion recoding, Private teachers Basal ganglia -(-)- Massive funnelling Tonic output firing Tectum Olfactory bulb Spinal cord Computational paradigms 100’s Myrs old that we fail to understand

















