Download

1 / 20
230 likes | 495 Views
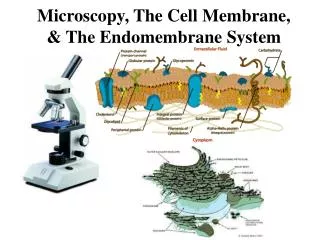
Endomembrane System. Yasir Waheed. NUST Center of Virology & Immunolgy National University of Sciences &Technology. The endomembrane system is composed of different membranes which are suspended in the cytoplasm of a eukaryotic cell.

E N D
Endomembrane System Yasir Waheed NUST Center of Virology & Immunolgy National University of Sciences &Technology
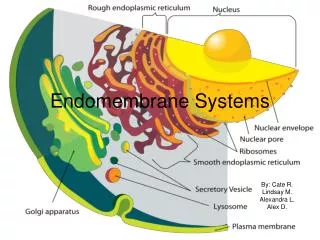
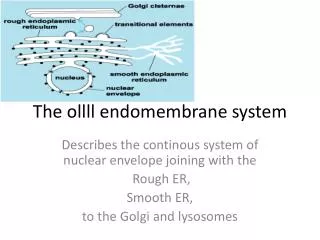
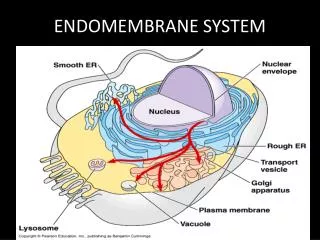
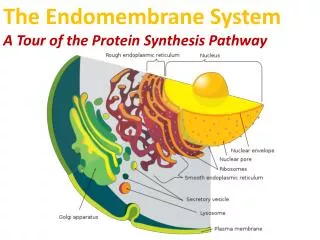
The endomembrane system is composed of different membranes which are suspended in the cytoplasm of a eukaryotic cell. These membranes divide the cell into functional and structural compartments or organelles. In eukaryotes the organelles of the endomembrane system include: the nuclear envelope, endopalsmic reticulum. etc
The Transport of Molecules between the Nucleus and the Cytosol • The nuclear envelope encloses the DNA and defines the nuclear compartment. This envelope consists of two concentric membranes that are penetrated by nuclear pore complexes. • Although the inner and outer nuclear membranes are continuous, they maintain distinct protein compositions. • The inner nuclear membrane contains specific proteins that act as binding sites for chromatin and for the protein of the nuclear lamina that provides structural support for this membrane. • The outer membrane is continuous with ER, where protein synthesis takes place.
Figure 12-9. The nuclear envelope. The double-membrane envelope is penetrated by nuclear pore complexes and is continuous with the endoplasmic reticulum. The nuclear lamina is a fibrous meshwork underlying the inner membrane.
Nuclear Pore Complexes Perforate the Nuclear Envelope • Nuclear envelope of all eukaryotes contains nuclear pore complexes, which is composed of more than 50 different proteins, called nucleoporins, that are arranged in octagonal symmetry . • The more active the nucleus is in transcription, the greater the number of pore complexes in its envelope. The nuclear envelope of a typicalmammaliancellcontains 3000 4000 pore complexes.
Nuclear pore complex contains four structural building blocks: column subunits, which form the bulk of the pore wall; annular subunits, which extend "spokes" (not shown) toward the center of the pore; lumenal subunits, which contain transmembrane proteins that anchor the complex to the nuclear membrane; and ring subunits, which form the cytosolic and nuclear faces of the complex. In addition, fibrils protrude from both the cytosolic and the nuclear sides of the complex. On the nuclear side, the fibrils converge to form basketlike structures.
Nuclear Import Export The compartmentalization of Ran-GDP and Ran-GTP. Localization of Ran-GDP to the cytosol and Ran-GTP to the nucleus results from the localization of two Ran regulatory proteins: Ran GTPase-activating protein (Ran-GAP) is located in the cytosol and Ran guanine nucleotide exchange factor (Ran-GEF) is bound to chromatin and is hence exclusively found in the nucleus.
Figure 12-16. A model for how GTP hydrolysis by Ran provides directionality for nuclear transport.
Figure 12-24. Three protein translocators in the mitochondrial membranes. The TOM and TIM complexes and the OXA complex are multimeric membrane protein assemblies that catalyze protein transport across mitochondrial membranes. The protein components of the TIM22 and TIM23 complexes that line the import channel are structurally related, suggesting a common evolutionary origin of both TIM complexes. As indicated, one of the core components of the TIM23 complex contains a hydrophobic alpha helical extension that is inserted into the outer mitochondrial membrane; the complex is therefore unusual in that it simultaneously spans two membranes.
Figure 12-29. Protein import from the cytosol into the inner mitochondrial membrane or intermembrane space. (A) A pathway that requires two signal sequences and two translocation events is thought to be used to move some proteins from the cytosol to the inner membrane. The precursor protein is first imported into the matrix space. Cleavage of the signal sequence (red) used for the initial translocation, however, unmasks an adjacent hydrophobic signal sequence (orange) at the new N terminus. This signal then directs the protein into the inner membrane, presumably by the same OXA dependent pathway that is used to insert proteins encoded by the mitochondrial genome.
(B) In some cases, the hydrophobic sequence that follows the matrix targeting signal binds to the TIM23 translocator in the inner membrane and stops translocation. The remainder of the protein is then pulled into the intermembrane space through the TOM translocator in the outer membrane, and the hydrophobic sequence is released into the inner membrane.
(C) Some soluble proteins of the intermembrane space may also use the pathways shown in (A) and (B) before they are released into the intermembrane space by a second signal peptidase, which has its active site in the intermembrane space and removes the hydrophobic signal sequence. (D) The import pathway used to insert proteins into the inner mitochondrial membrane utilizes the TIM22 complex, which is specialized for the translocation of multipass membrane proteins.
Figure 12-30. Translocation of a precursor protein into the thylakoid space of chloroplasts. (A) The precursor protein contains an N-terminal chloroplast signal sequence (red), followed immediately by a thylakoid signal sequence (orange). The chloroplast signal sequence initiates translocation into the stroma through a membrane contact site by a mechanism similar to that used for translocation into the mitochondrial matrix. The signal sequence is then cleaved off, unmasking the thylakoid signal sequence, which initiate translocation across thylakoid membrane.
(B) Translocation into the thylakoid space or thylakoid membrane can occur by any one of at least four routes: (1) a Sec pathway, so called because it uses components that are homologs of Sec proteins, which mediate protein translocation across the bacterial plasma membrane (2) an SRP-like pathway, so called because it uses a chloroplast homolog of the signal recognition particle, or SRP . (3) a pH pathway, so called because it is driven by the H+ gradient across the thylakoid membrane, and (4) a spontaneous insertion pathway that seems to require no protein translocator for membrane integration.
Figure 12-46. A model for how a soluble protein is translocated across the ER membrane. On binding an ER signal sequence (which acts as a start-transfer signal), the translocator opens its pore, allowing the transfer of the polypeptide chain across the lipid bilayer as a loop. After the protein has been completely translocated, the pore closes, but the translocator now opens laterally within the lipid bilayer, allowing the hydrophobic signal sequence to diffuse into the bilayer, where it is rapidly degraded.
Transmembrane, Rhodopsin Figure 12-47. How a single-pass transmembrane protein with a cleaved ER signal sequence is integrated into the ER membrane. In this hypothetical protein the cotranslational translocation process is initiated by an N-terminal ER signal sequence (red) that functions as a start-transfer signal. In addition to this start-transfer sequence, however, the protein also contains a stop-transfer sequence (orange). When the stop-transfer sequence enters the translocator and interacts with a binding site, the translocator changes its conformation and discharges the protein laterally into the lipid bilayer.