Download
1 / 1
10 likes | 301 Views
Exploiting Protein Cage Dynamics for Engineering Active Nanostructure s NSF NIRT CBET-0709358 Brian Bothner ad , Yves Idzerda bd , Mark Young cd , Trevor Douglas ad (PI)

E N D
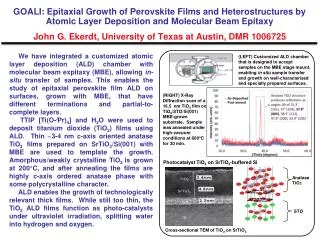
Exploiting Protein Cage Dynamics for Engineering Active NanostructuresNSF NIRT CBET-0709358Brian Bothnerad, Yves Idzerdabd, Mark Youngcd, Trevor Douglasad (PI) a.Department of Chemistry and Biochemistry, b. Department of Physics, c. Department of Plant Sciences, d. Center for Bio-Inspired Nanomaterials, Montana State University, Bozeman, MT 59717 Fe2+ Room Temperature Empty PfFn Heat Treatment at 100°C swollen 31.5 nm Shear 4.2 MPa Young’s 10.2 MPa < 1 sec. Objectives: The development of protein cage architectures as dynamic size and shape constrained templates for nanomaterials synthesis using three integrated components - Active Interfaces, Cage Dynamics, and Confinement. closed 28.6 nm Shear 9.8 MPa Young’s 24.7 MPa Internally Constrained Proteins for Catalyst Scaffolds TiO2 in CCMV with Photocatalytic Activity Polymer in Hsp – increased thermal stability Nanoparticles of TiO2 were grown inside the CCMV capsid by polymerization of a Ti-oxo anion (Ti-BALDHI) at high pH. Pair Distribution Function (PDF) analysis showed a structure similar to beta-TiO2, with a crystalline domain size of 1.3nm. This is significantly smaller than the particle sizes shown by TEM, and indicates that each protein cage likely contains numerous small crystallites, rather than a single TiO2 crystal. The virus-templated TiO2 nanoparticles have been shown to by photochemically active, and may be useful for light-harvesting applications. A cross-linked branched polymer can be initiated and synthesized within the interior cavity of a protein cage architecture. This polymer network allows for the spatial control of pendant reactive sites and dramatically increases the stability of the cage architecture extending the thermal stability of the protein cage to at least 120 °C. The polymer was generated sequentially coupling multifunctional monomers using click chemistry to create a branched cross-linked polymer network through generation 2.5 where it was limited by the size of the cavity. A range of protein cage architectures can be used as synthetic templates The interior, exterior, and subunit interfaces can be chemically and genetically modified. Protein cage library size range from 30 to 10 nm diameter. CCMV (30nm), Ferritin (12 nm), sHsp (12 nm), Dps (10 nm) Generations 2.5 exhibited exceptional stability upon exposure to heat. The G41C native cage decomposes upon exposure to 120 °C for 10 min while generation 2.5 remains intact upon exposure to 120 °C for 30 min. TEM of unstained (right) and uranyl acetate stained (left) CCMV reacted with Ti-BALDHI. The diffraction pattern (insert) could be indexed to b-TiO2. The scale bars are 100 nm. Coordination polymer in Hsp a-Fe2O3 in Pf_Fn with Photocatalytic Activity A branched iron-phenanthroline based coordination polymer has been constructed in a water based system using a click chemistry approach to link monomeric coordination complexes together within a protein cage nano-architecture, which acts both a template and a sized constrained reaction environment. Using ferritin from the hyperthermophile Pyrococcus furiosis we have mineralized a disorodered Fe(O)OH within the protein cage and transformed it to hematite at 100ºC in the presence of oxalate. The protein remains intact during this process. The hematite mineral acts as a semiconductor with a bandgap in the visible. Coomassie stained acid urea denaturing gel evidence of cross-linking of the protein subunits. As the generations increase across the gel, the monomer, which is dominant in G0.0 and G0.5, gradually decreases with a parallel increase in intensity at the top of the gel. The ratio of the absorbance at 530 to 280 nm at each generation is related to the number of Fe(II) moieties per a cage. The absorbance at 530 nm is only due to the Fe(az-phen)32+, while the absorbance at 280 nm is from both the protein and the Fe(az-phen)32+. Cartoon schematic for the conversion of a disordered iron oxide to an ordered photoactive iron oxide. Photocurrents generated by the β-TiO2 / CCMV composites upon exposure to a high intensity LED (420 nm) or UV (365 nm) light source. The response to the visible illumination is negligible. Experimental PDF compared with β-TiO2 and NaCl phases. A good fit was obtained through a linear combination of the two components. The stepwise formation of a coordination polymer inside Hsp G41C starts at the internal cysteine of each protein subunit. Each generation is added through the introduction of either an alkyne or azide compound. Rapid and Reversible Environmental Response The development of rotein cages that can react to environmental signals leading to changes in confomation, stability, and chemical potential are being investigated Redox active switching of DPS cages. C101S WT flexible rigid DPS swollen Noncovalent dye binding shows that reduction of the disulfide in WT DPS decreases cage thermal stability. A mutant form lacking the disulfide is insensitive to redox potential. O = oxidized, R = reduced. frequency closed Transmission electron micrographs of (A) unstained and (B) stained refluxed Pf-Fn. The dark electron dense cores in (A) have an electron diffraction pattern (insert) consistent with -Fe2O3. Photocurrent generated upon exposure of hematite in Pf_Fn with methyl viologen (blue dash-dotted), ferrihydrite in Pf_Fn (red dashed), and methyl viologen (black solid) to a high intensity LED light. time Quartz crystal microbalance analysis of the closed to swollen transition. Modulus of CCMV protein cages reversibly transitions between two states, controlled with pH and divalent cations. 40 50 60 70 80 90 C