Download
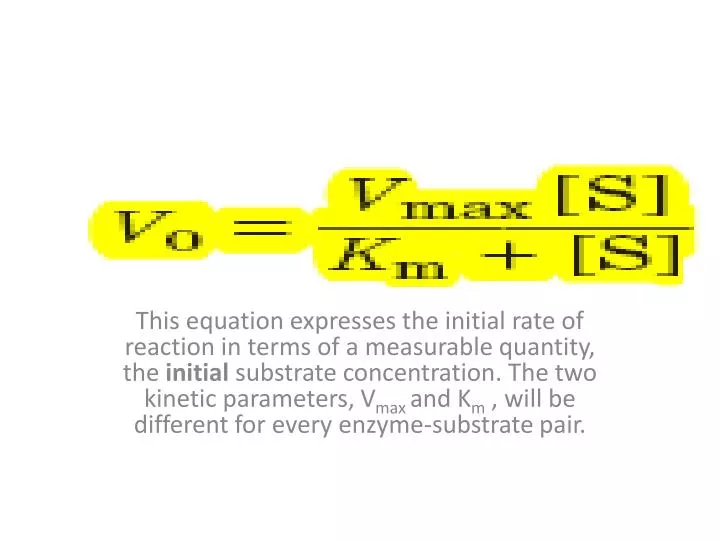
1 / 31
490 likes | 1.18k Views
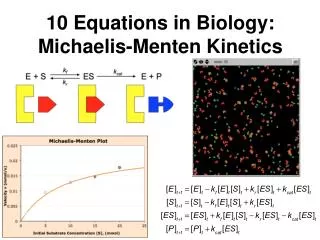
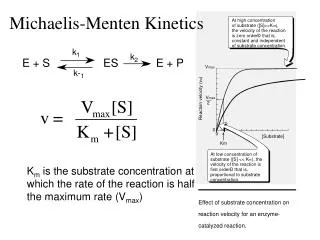
This equation expresses the initial rate of reaction in terms of a measurable quantity, the initial substrate concentration. The two kinetic parameters, V max and K m , will be different for every enzyme-substrate pair. . When [S] >> K m. When [S] << K m.

E N D
This equation expresses the initial rate of reaction in terms of a measurable quantity, the initial substrate concentration. The two kinetic parameters, Vmaxand Km , will be different for every enzyme-substrate pair.
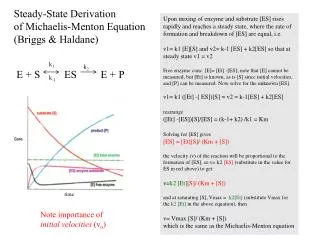
When [S] >> Km When [S] << Km The Michaelis-Menten Equation nicely describes the experimental observations. The substrate concentration at which V0 is half maximal isKm
The Vmax and Km values of a certain enzyme can be measured by the double reciprocal plot (i.e., the Lineweaver-Burk plot).
The double reciprocal plot: 1/V0 vs 1/[S]
The Michaelis-Menten equation, but not their approximated mechanism applies to a great many enzymes • Most enzymes (except the regulatory enzymes) have been found to follow the Michaelis-Menten kinetics, but their actual mechanisms are usually more complicated (by having more intermediate steps) than the one assumed by Michaelis and menten. • The values of Vmax and Km alone provide little information about the number, rates, or chemical nature of discrete steps in the reaction.
Vmax is determined by kcat, the rate constant of the rate-limiting step • Vmax = kcat[Et] • kcatequals to k2 or k3 or a complex function of both, depending on which is the rate-limiting step. • kcat is also called the turnover number: the number of substrate molecules converted to product in a given unit of time per enzyme molecule when the enzyme is saturated with substrate.
40,000,000 molecules of H2O2 are converted to H2O and O2 by one catalase molecule within one second!
The kinetic parameters kcat and Km are often studied and compared for different enzymes • Km often reflects the normal substrate concentration present in vivo for a certain enzyme. • The catalytic efficiency of different enzymes is often compared by comparing their kcat/Km ratios (the specificity constant). • when • [S]<<Km • kcat/Km is an apparent second-order rate constant (with units of M-1S-1), relating the reaction rate to the concentrations of free enzyme and substrate.
The value of kcat/Km has an upper limit (for the perfected enzymes) • It can be no greater than k1. • The decomposition of ES to E + P can occur no more frequently that E and S come together to form ES. • The most efficient enzymes have kcat/Km values near the diffusion-controlled limit of 108 to 109 M-1S-1.
Catalytic perfection (rate of reaction being diffusion-controlled) can be achieved by a combination of different values of kcat and Km.
Rates of individual steps for an enzyme-catalyzed reaction may be obtained by pre-steady state kinetics • The enzyme (of large amount) is used in substrate quantities and the events on the enzyme are directly observed. • Rates of many reaction steps may be measured independently. • Very rapid mixing and sampling techniques are required (the enzyme and substrate have to be brought together in milliseconds and measurements also be made within short period of time).
“Rapid kinetics” or “pre-steady- state kinetics”is applied to the observation of rates of systems that occur in very short time intervals (usually ms or sub-ms scale) and very low product concentrations. This period covers the time from the enzyme encountering its target (either a substrate, inhibitor or some other ligands) to the point of system settling to equilibrium. The concentration of ES will rise from zero to its steady-state value. (ms or sub-ms)
Solutions are forced together very rapidly. Stopped-flow apparatus for pre-steady state kinetics (since 1940s)
Quench flow apparatus for rapid kinetics
Cofactors and Coenzymes • Some enzymes do not need any additional components to show full activity. • However, others require non-protein molecules called cofactors to be bound for activity. • Cofactors can be either inorganic (e.g., metal ions and iron-sulfur clusters) or organic compounds, (e.g., flavin and heme).
Cofactors • Active enzyme / Holoenzyme: • Polypeptide portion of enzyme (apoenzyme) • Nonproteineous group (cofactor) • Cofactors are bound to the enzyme for it to maintain the correct configuration of the active site • Metal ions • Organic compounds • Organometallic compounds
Organic cofactors can be either: • prosthetic groups, which are tightly bound to an enzyme, or coenzymes, which are released from the enzyme's active site during the reaction. • carbonic anhydrase, with a zinc cofactor bound as part of its active site. These tightly-bound molecules are usually found in the active site and are involved in catalysis.
For example, flavin and heme cofactors are often involved in redoxreactions. Most cofactors are not covalently attached to an enzyme, but are very tightly bound. However, organic prosthetic groups can be covalently bound.
Coenzymes Coenzymes are small organic molecules that transport chemical groups in an enzymatic reaction. OR Any of a number of freely diffusing organic compounds that function as cofactors with enzymes in promoting a variety of metabolic reactions. Some of these chemicals such as riboflavin, thiamine and folic acid are vitamins, (acquired).
The chemical groups carried include the hydride ion (H-) carried by NAD or NADP+, the acetyl group carried by coenzyme A, … etc. • Since coenzymes are chemically changed as a consequence of enzyme action, it is useful to consider coenzymes to be a special class of substrates, or second substrates, which are common to many different enzymes. • About 700 enzymes are known to use the coenzyme NADH.
Coenzymes are usually regenerated and their concentrations maintained at a steady level inside the cell: for example, NADPH is regenerated through the pentose phosphate pathway and S-adenosylmethioninebymethionineadenosyltransferase • Coenzymes participate in enzyme-mediated catalysis in stoichiometric (mole-for-mole) amounts and are modified during the reaction.
Coenzymes • A coenzyme is required by some enzymes • An organic molecule bound to the enzyme by weak interactions / Hydrogen bonds • Most coenzymes carry electrons or small groups • Many have modified vitamins in their structure
Nicotinamide Adenine Dinucleotide in Oxidized and Reduced Forms 19.7 Cofactors and Coenzymes
NAD+ to NADH Mechanism • The nicotinamide part of NAD+ accepts a hydride ion (H plus two electrons) from the alcohol to be oxidized • The alcohol loses a proton (H+ ) to the solvent 19.7 Cofactors and Coenzymes +H+ Oxidized form Reduced form
Two Other Adenine Dinucleotide Coenzymes 19.7 Cofactors and Coenzymes
Enzymes • Papainsadiaarshad • Bromelainridarehman • Trypsin, Chymotrypsin • Pancreatinrafia • Pepsin • Diastase tahira • Pectinaseamna • Urokinaseshamyam • Asparaginaseaimen • Hyaluronidase AYSHA nAEEM • N-acetyl-galactosamineAYSHA SAJID • DNA-aseRabiyatahir
Kinase AYSHA zaheer • Phosphataseaqsa • RDRP fatima • Topoisomeraseabida • Lactase hafza • Amylase saba • NO synthaseazka • Luciferaseadeena • EC1 Badar and Uzair • DDPoleymeraseBakhtawar