Download
1 / 47
480 likes | 654 Views
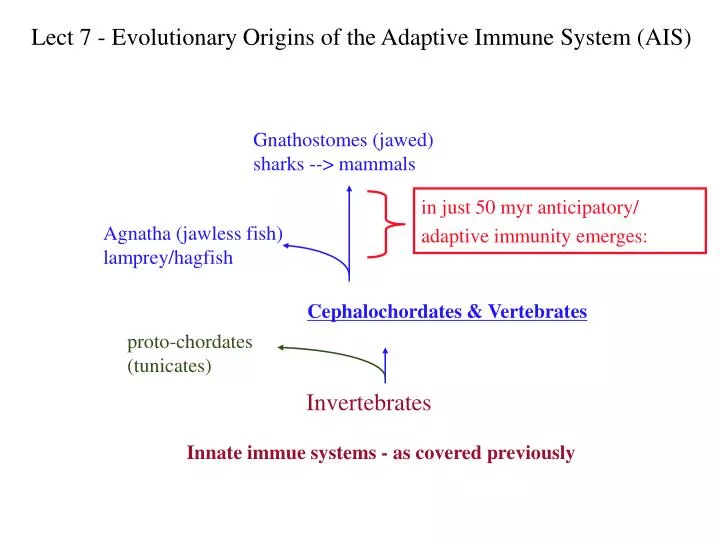
Lect 7 - Evolutionary Origins of the Adaptive Immune System (AIS). Gnathostomes (jawed) sharks --> mammals. in just 50 myr anticipatory/ adaptive immunity emerges:. Agnatha (jawless fish) lamprey/hagfish. Cephalochordates & Vertebrates. proto-chordates (tunicates). Invertebrates.

E N D
Lect 7 - Evolutionary Origins of the Adaptive Immune System (AIS) Gnathostomes (jawed) sharks --> mammals in just 50 myr anticipatory/ adaptive immunity emerges: Agnatha (jawless fish) lamprey/hagfish Cephalochordates & Vertebrates proto-chordates (tunicates) Invertebrates Innate immue systems - as covered previously
How could you determine the evolutionary origins of the AIS? first you need to establish the endpoints that you’re looking for the evolutionary origins of – i.e. what are the key elements of AIS?
Only recently it was established that Agnathans have an AIS system that is analogous to our AIS system - with cells with B- and T-cell like features – next lecture. Until then most work on origins of AIS focused on molecular evolution Gnathostomes (jawed) sharks --> mammals unique Adaptive Immune System emergence of AIS Agnatha (jawless fish) lamprey/hagfish Cephalochordates & Vertebrates proto-chordates (tunicates) Invertebrates
How could you determine the evolutionary origins of the AIS? From what organisms are you establishing your endpoints? think of the most parsimonious (simplistic) system – where would you find this (to establish AIS endpoints)?
Throw-backs and relicts?! V D J C V J C Ig heterodimer Ig homodimer
How or where do you identify adaptive immunity precursors? Presumptions – that precursor was retained in some organism & that organisms retain their ancestral form – i.e. that the modern animal is a fixed-in-time version of when evolution diverged from it Are these necessarily true? sometimes …. e.g. human fetal gill slits Two scenarios for where to look for precursor precursor changed function and evolved into its’ AIS function (think tachylectin to fibrinogen) you’d look in ? precursor gene was duplicated and one copy retained precursor structure (think C3, C4 & C5) you’d look in ?
How do you piece together the evolutionary path of cells or molecules? Identify the ‘new’ traits that you are looking for – i.e. the endpoint Attempt to identify the precursors based on structure &/or function Attempt to identify any possible contributing factors
What was new about gnathostome adaptive immunity? 1) New type of Ig domain - C1 – subtle changes in protein organization 2) A split V-type Ig domain - i.e. V-, D-, & J-elements ….. 3) …. and the VDJ recombination mechanism to join the elements 4) The peptide binding groove of MHC for Ag presentation 5) Lymphocytes & lymphoid organs
Events that may have contributed to the evolution of adaptive immunity: 1) Emergence of V1 type and C1 type Ig domains (? cause or conseq) 2) Two whole Genome duplications in early vertebrates *3) Invasion of the transposons - splitting & recombining V domains - into V & J-elements (and later also D-element) * Hypothesis
Where to look first? Ig domains exist in all eukaryotes and are just a pair of folded-over b-pleated sheets The origins of the MHC-fold are less clear Klein and Nikolaidis (2005) Proc. Natl. Acad. Sci. USA 102, 169-174
Changes to the Ig domains are associated with AIS evolution Variable & Constant immunoglobulin domains exist in many molecules and in many forms: Variable, C1 or C2-Constant & Intermediate type Ig domains Those in the TcR and Ig molecules have some distinct features & defining those features provides the framework for finding the ancestral Ig/TcR molecule Though the distinctions in protein traits can be quite subtle ....
Ig superfamily (IgSF) members from throughout the animal kingdom are of a particular V and C2 type These V-domains are encoded by a single exon that is not the product of VDJ rearrangement V V V V V V V V V Kuby 4-19
The key proteins of adaptive immunity have C1-type Ig domains C1 domains only found in proteins of adaptive immunity? (Ig, TcR, MHC & tapasin) & the variable domain of the TcR & BCR is encoded on an ‘assembled’ VDJ exon V V V V V V What defines the Ig domain subsets? Kuby 4-19
Ig L-chain C1-domain & V-domain Kuby 4-6 Ig domain types are defined by: # b-strands, type ofb-strand interactions & location of -S-S- (disulfide) bond (black bar) 4 3 4 5 D E B A G F C D E B A G F C C' C" C1-type
V-set Ig domain vs. I-set Ig domain V-set purple is-strand that doesn't contribute to Ig fold Intermediate-set http://pps98.man.poznan.pl/assignment/projects/ruiz/PROJ/HTML1.html
Comparison of C1- and C2-set Ig domains C1 C2 http://pps98.man.poznan.pl/assignment/projects/ruiz/PROJ/HTML1.html
Comparison of V- and C2-set Ig domains corresponds to J-element region C2 http://pps98.man.poznan.pl/assignment/projects/ruiz/PROJ/HTML1.html
What would an ancestral Ig/TcR look like? An immune function!? V-type and C1-type(?) domains V-domain with a diglycine bulge in b-strand ‘G’ (J-element like) Transmembrane & cytoplasmic domains - receptor signaling?
Some vertebrate proteins - V domains with a fused J-element like region (there’s no recombination – one exon encoding V & J-like elements) true J-elements encode a "diglycine bulge" (F)GxGTxLxV In Ig & TcR the diglycine bulge facilitates dimer formation between VL & VH (in Ig) or between Va & Vb (of TcR) - it also pushes hypervariable loops outward towards ligand The ancestral gene to Ig & TcR is expected to have this trait ......
C1? TM TM Cyto Cyto C2 C2 VJ VJ Pre-ancestral approach - survey extant chordates lacking the AIS Tunicate - Ciona genome (protochordate - larval notochord) Two candidate predecessors to Ig and TcR in Ciona genome .... 1) Nectin - cell adhension molecules (& poliovirus receptor) 2) CTX/ JAM - cell junctionaladhesion molecules - also found on cortical thymocytes (originally in Xenopus) DuPasquier 2004 C.R. Biologies 327: 591-601
VJ/C2 VJ/C2 VJ VJ ITIM ITAM 2nd approach - assume that ancestral TcR/Ig has been retained on copy e.g. CTX/JAM - as before - but in higher vertebrates too. or NITR - novel immune type receptor - in fish (mammalian homologs too) have a V- and I-type domain & is membrane bound they’re multigenic and polymorphic - early allorecognition function? have ITIM or ITAM signaling tails in cytoplasm! NITRs Trends in Immunology 25(1):11-16
BCR & TcR have associated ITAM signaling domains NK receptors & others have ITIM inhibitory domains NITRs were the best candidate common ancestor to Ig/TcR Trends in Immunology 25(1):11-16 * - split VJ
signal regulatory proteins (SIRPs) also resemble predicted ancestor Identified in mammals & birds - thus far have a fused V-J & 2 C1-type domains and have direct or indirect signaling domains they’re a cluster of genes - along with NK genes Regulate phagocytosis & ? Proposed original function: clearing self cells that are damaged Trends in Immunology 25(1):11-16
Whole genome duplication - another major evolutionary event Provided duplicate or spare copies of essential genes Duplicates can be modified without altering function of originals
Duplication of genes provides 'spare' copies Chromosome Duplication Paralogous Chromosomes Time 2nd Duplication 4 copies of each gene
Spare genes can be altered: Modification of original gene Chromosome Duplication Time 1 1 Duplication
Spare genes can be altered: Modification of original gene Complete loss of copy Chromosome Duplication Time 1 2 1 Duplication
Spare genes can be altered: Modification of original gene Complete Loss Intrachromosomal duplications Chromosome Duplication Time 1 3 2 1 Duplication
Spare genes can be altered: Modification of original gene Complete Loss Intrachromosomal duplications Pseudogenes () - Incomplete loss Chromosome Duplication Time 1 3 2 1 Duplication 4
There is evidence that 2 genome duplication events took place early in vertebrate evolution - at the time that adaptive immunity emerged (Flajnik & Kasahara. 2002 Immunity, 15:351-362; Nat. Rev. Genetics 2010, 11:47-59)
Attempts to identify early MHC by Synteny Analysis led to the evidence that the ‘whole genome duplication hypothesis’ was correct Premise of synteny analysis: If in gnathostomes: chromosome Z housekeeping gene Q housekeeping gene R MHC class I housekeeping gene S housekeeping gene T MHC class II housekeeping gene U housekeeping gene V Then in pre-gnathostomes: chromosome Z equivalent housekeeping gene Q housekeeping gene R pre-MHC? housekeeping gene S housekeeping gene T pre-MHC? housekeeping gene U housekeeping gene V
Synteny caveat – chromosomes can change by translocations etc. mouse Chr 11 shares synteny with 6 diff. human chromosomes PLoS Genet. 2009 5(12): e1000759.
Evidence of paralogous (duplicated) chromosomes in mammals: The same paralogous genes are found on the same chromosomes Human Chr 6Chr 9Chr 1Chr 19 B3GALT4 B3GALT2 B3GALT RXRB RXRA RXRG COL11A2 COL5A1 COL11A1 Ring3 Ring3L BRDT HUNK1 Notch4 Notch1 Notch2 Notch3 PBX2 PBX3 PBX1 C4 C5 C3 Bf C2 HSP70 HSPA5 HSPA6 MICA CD1 MICB CD1 NTRK4 NTRK2 NTRK1 some genes are lost, new or duplicated – chromosomes have evolved
Notice that complement genes were duplicated within chromosomes or when the chromosomes duplicated Human Chr 6Chr 9Chr 1Chr 19 B3GALT4 B3GALT2 B3GALT RXRB RXRA RXRG COL11A2 COL5A1 COL11A1 Ring3 Ring3L BRDT HUNK1 Notch4 Notch1 Notch2 Notch3 PBX2 PBX3 PBX1 C4 C5 C3 Bf lost? C2 HSP70 HSPA5 HSPA6 MICA CD1 MICB CD1 NTRK4 NTRK2 NTRK1
Also notice that related non-classical MHC genes are on 2 chromosomes so they evolved before the second whole genome duplication Chr 6Chr 9Chr 1Chr 19 B3GALT4 B3GALT2 B3GALT RXRB RXRA RXRG COL11A2 COL5A1 COL11A1 Ring3 Ring3L BRDT HUNK1 Notch4 Notch1 Notch2 Notch3 PBX2 PBX3 PBX1 C4 C5 C3 Bf C2 HSP70 HSPA5 HSPA6 MICA CD1 MICB CD1 NTRK4 NTRK2 NTRK1
While classical MHCs are just on one chromosome – consistent with their emergence after the second genome duplication - post-agnatha Human Chr 6Chr 9Chr 1Chr 19 B3GALT4 B3GALT2 B3GALT RXRB RXRA RXRG COL11A2 COL5A1 COL11A1 Ring3 Ring3L BRDT HUNK1 Notch4 Notch1 Notch2 Notch3 PBX2 PBX3 PBX1 C4 C5 C3 Bf C2 HSP70 HSPA5 HSPA6 MICA CD1 MICB CD1 NTRK4 NTRK2 NTRK1 classical MHC class II genes MHC class III genes classical MHC class I genes
V J Hypothesis for evolutionary origin of diversity in antibody receptors SIRP or nectin-like gene? V C (1) transposon insertion transposon inserts and splits the exon (2) Ancestor gene to IgL-chain TcRa and TcRg C (3) transposon inserts again --> V-, D- & J-elements of IgH, TcRb & TcRd
recombination signal sequences inverted repeated VDJ Recombination vs. Transposition U U Transposons encode and use a transposase to excise & insert their gene
Were RAG genes derived from a transposon? Early research failed to find sequence homologies between RAG and known transposases but other similarities were noted Similar chemistry of DNA targeting by excision/insertion enzyme: - RAG recognizes a specific sequence motif - RSS - transposase recognizes specific sequences - Inverted Repeats - both have free hydroxl attack of phosphodiester bond - requires Mg++/Ca++ but not ATP - some Transposons form a hairpin loop after removing self from host
Other important functional similarities between RAG & Transposons RAG & transposase stay attached to DNA following excision RAG recombinases can operate as transposons in vitro - i.e. the excised DNA can be re-inserted back into DNA too (instead of forming the signal joint) In 2004 a new class of Transposon was recognized & the core catalytic site of the Transibtransposase shared homology with RAG2 core
Transib – shares homology with RAG1 www.plosbiology.org/article/info:doi/10.1371/journal.pbio.0030181 Transib’s RAG1s shared color equals shared amino acids Kapitonov and Jurka Plos Biology 2005 RAG1 Core and V(D)J Recombination Signal Sequences Were Derived from Transib Transposons
The inverted repeats (IRs) of Transib transposons also show homology with the RAG targets – Recombination Signal Sequences (RSSs) Transib IRs are short conserved sequences (~10 to 14 nt) separated by variant nucleotides of ~ 5 to 23 bp in length Some of the conserved nucleotides are similar to the RSS heptamer RSSs are conserved heptamer (7 nt) and nonamer (9 nt) separated by variant nucleotides of either 12 or 23 bp (+/- 1 bp) in length Kapitonov and Jurka. Plos Biology 2005 RAG1 Core and V(D)J Recombination Signal Sequences Were Derived from Transib Transposons
RAG1-like first shows up in pre-chordate deuterostomes, while RAG2 arises only in gnathostomes (?) Kapitonov and Jurka; Plos Biology 2005 RAG1 Core and V(D)J Recombination Signal Sequences Were Derived from Transib Transposons
Proposal for how the primitive RAG genes split the Variable domain exon into V, D & J – the IRs are left in the Ig gene while the RAG precursors move elsewhere in the genome (unknown) Fugmann et. al. Proc. Natl. Acad. Sci. 2006, 103 (10): 3728-3733
FYI - Defining relationships between genes (structural) (http://www.ncbi.nih.gov/Education/BLASTinfo/Orthology.html)