Download

1 / 23
230 likes | 430 Views
( Kellen L. Olszewski,Michael W. Mather,Joanne M. Morrisey,Benjamin A. Garcia,Akhil B. Vaidya,Joshua D. Rabinowitz,and Manuel Llinás ). BRANCHED TRICARBOXYLIC ACID METABOLISM IN Plasmodium falciparum. Submitted by Selma Abdul Samad BCH 10-05-02 S2 MSc.Biochemistry

E N D
( Kellen L. Olszewski,Michael W. Mather,Joanne M. Morrisey,Benjamin A. Garcia,Akhil B. Vaidya,Joshua D. Rabinowitz,and Manuel Llinás ) BRANCHED TRICARBOXYLIC ACID METABOLISM IN Plasmodium falciparum Submitted by Selma Abdul Samad BCH 10-05-02 S2 MSc.Biochemistry Dept. of Biochemistry
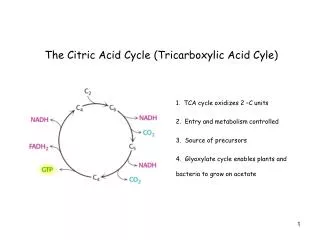
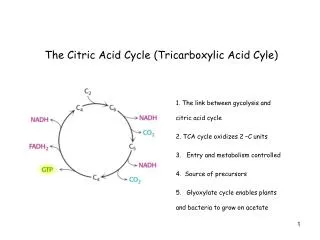
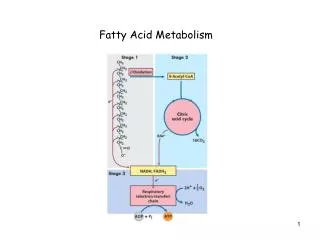
TCA cycle • Also called citric acid cycle • The central hub of carbon metabolism – connects glycolysis,gluconeogenesis,respiration, aminoacid synthesis and other biosynthetic pathways • Final products – CO2 , water and energy • Begins with the transfer of a two-carbon acetyl group from acetyl-CoA to the four-carbon acceptor compound (oxaloacetate) to form a six-carbon compound (citrate) • The citrate then goes through a series of chemical transformations, losing two carboxyl groups as CO2. The carbons lost as CO2 originate from what was oxaloacetate, not directly from acetyl-CoA
For each acetyl group that enters the citric acid cycle, three molecules of NADH are produced • At the end of each cycle, the four-carbon oxaloacetate has been regenerated, and the cycle continues • Several catabolic pathways converge on the TCA cycle • The citric acid cycle is regulated by the energy needs of the cell • So,ATP and NADH exert a negative feed back control on citrate synthase and iso-citrate dehydrogenase
Plasmodium falciparum • A protozoan parasite • cause malaria in humans • transmitted by the female Anopheles mosquito • Most dangerous of the Plasmodium species • It causes severe malaria via a distinctive property not shared by any other human malaria, that of sequestration.Withinthe blood stage cycle, the mature forms change the surface properties of infected erythrocytes causing them to stick to endothelial walls of blood vessels(a process called cytoadherence).This leads to obstruction of the microcirculation and results in dysfunction of multiple organs, typically the brain in cerebral malaria.
The life cycle of all Plasmodium species is complex • Infected female Anopheles mosquito bites Sporozoites released from its salivary gland enters bloodstream and invade hepatocytes For 14 days liver-stage parasites differentiate ;undergo asexual multiplication forming merozoites Merozoites invade erythrocytes ; continue multiplication ; invade more and more erythrocytes Malaria with all its symptoms manifested(fever ,chill, RBC rupture)
TCA metabolism inPlasmodium falciparum • Blood-stage Plasmodium parasites rely almost entirely on glucose fermentation for energy and consume minimal amounts of oxygen • Yet the parasite genome encodes all of the enzymes necessary for a complete TCA cycle • By tracing 13C-labeled compounds using mass spectrometry,the researchers showed that TCA metabolism in the human malaria parasite P. falciparum is largely disconnected from glycolysis and is organized along a fundamentally different architecture • This pathway is not cyclic but rather a branched structure in which the major carbon sources are the amino acids glutamate and glutamine
As a consequence of this branched architecture, several reactions must run in the reverse of the standard direction thereby generating two-carbon units in the form acetyl-CoA • It is further shown that glutamine-derived acetyl-CoA is used for histoneacetylation while glucose-derived acetyl-CoA is used to acetylateaminosugars • Thus the parasite has evolved two independent acetyl-CoA-production mechanisms with different biological functions
Metabolites in red – wastes to be effluxed into medium Red arrows – reductive pathway of TCA metabolism Blue arrows – oxidative pathway * - citrate cleavage step enzyme (unclear) ** - 2 enzymes are predicted – cytosolicmalatedH or mitoch. malate:quinoneoxidoreductase OG – 2-oxoglutarate ; ICT – Isocitrate ; Ac-R – Acetate/acetyl Co.A
The mitochondrion of P.falciparum - contains smallest genome sequenced - evolved reduced functional roles - limited mitochondrial cristae - minimal oxygen consumption - rapid glucose fermentation All these suggest that oxidative phosphorylation is not a significant source of ATP generation during the blood stage • The critical metabolic function of electron transport during blood stage growth is the regeneration of ubiquinone in order to supply pyrimidine biosynthesis • TCA metabolism plays an active role in the metabolism of the parasite. The parasite genome encodes orthologues for all TCA cycle enzymes, which are all transcribed during the blood stage
The presence of an essential de novo heme biosynthesis pathway in P. falciparum further implies that succinyl-CoA must be generated in the mitochondrion • It has recently been demonstrated that the P. falciparum PDH complex localizes not to the mitochondrion but the apicoplast • Thus,instead of its canonical role of feeding glucose-derived carbon into the TCA cycle, the suggested role of PDH is solely to produce acetyl-CoA for fatty acid elongation • The major carbon source contributing to the accumulation of TCA intermediates was determined - By culturing synchronized parasite-infected RBCs in media supplemented with C13 labelled glucose,C13 and N15 labelled aspartate & glutamine ( LC and MS) • In cultures with C13labelled glucose, all glycolytic intermediates were labelled, while carboxylic acid pool labelling was limited (some malate and fumarate )
The absence of labeling into other TCAintermediates suggests that these labeleddicarboxylic acids derive from cytosolic pathways independent of mitochondrial TCA metabolism • Similarly, growth on 13C-15N-aspartate results only in the generation of 13C-malate and 13C-fumarate which can also occur in the cytosol • Feeding of labeled glucose results in labeling of only a small fraction of the total acetyl-CoA pool, suggesting the presence of additional sources for two-carbon units • These data raised the possibility that glucose and aspartate-derived metabolites are disconnected from mitochondrial TCA metabolism • Fed with C13N15glutamine – significant labelling of all TCA compounds • Extracellular glutamine is rapidly taken up by parasitized RBCs and deamidated to glutamate, which can donate its carbon skeleton to TCA metabolism through conversion to 2-oxoglutarate
Figure Glutamine drives reverse flux through the TCA cycle
And finally it is concluded from the datas that several TCA cycle reactions are running with net flux in the reverse direction, in the process generating C2 units from 2-oxoglutarate via citrate • To further dissect the biological role of this reverse TCA branch,the major metabolic fates for C2 units: fatty acid synthesis, protein modification and small molecule acetylation,were investigated • When parasites are cultured in medium containing either 13Cglucose or 13C-15N-glutamine ,robust labeling of the acetyl groups was observed,inhistonetails,only in the 13C-15N-glutamine-fed cultures • However,UDP-GlcNAc,a nucleotide aminosugar acetylated in the endoplasmic reticulum during the biosynthesis of GPI-anchored proteins associated with malaria pathogenesis,islabeled at the acetyl group only during growth on 13C-glucose
Figure 2 Acetyl groups deriving from glucose and glutamine are functionally distinct
Thus it appears that the malaria parasite has evolved two independent pathways that produce acetyl-CoA for different metabolic functions • How glucose and glutamine derived C2 units are maintained as functionally distinct pools and transported from their respective organelles to different sites of acetylation remains to be investigated • The metabolic labeling data suggest a branched architecture for mitochondrial carbon metabolism in which both arms produce malate. In order to achieve a net flux through these pathways it would be necessary to remove this terminal product, either by conversion or excretion • They found that malate, 2-oxoglutarate and, to a lesser extent, fumarate are excreted from infected RBCs at a significant rate • Based on these results,a new model for central carbon metabolism in blood stage Plasmodium sp. was proposed
Figure Malate excretion by P. falciparum-infected RBC cultures
Figure An integrated model for central carbon metabolism in P. falciparum
In this pathway the ultimate carbon source for mitochondrial carboxylic acid pools are the amino acids glutamine and glutamate, and carbon flux in the mitochondrion is organized into two independent linear branches • Branch 1 begins with the reductive carboxylation of 2-oxoglutarate to isocitrate, which is then isomerized to citrate. This citrate is cleaved into a C2 compound and oxaloacetate, which is reduced to malate (red in figure) • Branch 2 comprises the standard clockwise turning of the TCA cycle to oxidize 2-oxoglutarate to malate, in the process generating reducing power and succinyl-CoA, an essential precursor for heme biosynthesis (blue in figure) • The fact that two labeled forms are observed for malate and fumarate but no other TCA intermediates during growth on 13C-15N-glutamine suggests that both branches converge at these metabolites and they are the terminal products of each
The P. falciparumgenome encodes only an NADP(H) specific, mitochondrial IDH, suggesting that it may have entirely lost the ability to run a TCA cycle and is effectively locked into this branched architecture • It is proposed that the mitochondrial NADPH required by this reductive pathway may be generated by the parasite's NADP(H)-specific glutamate dehydrogenase and glutamate oxidation has been detected in isolated P. falciparummitochondria • This branched TCA pathway maybe an evolutionary trade-off in which metabolic flexibility is lost in order to optimize growth within the specific environment of the host cell
Reference Nature. 2010 August 5; 466(7307): 774–778. Branched Tricarboxylic Acid Metabolism in Plasmodium falciparum Kellen L. Olszewski,1 Michael W. Mather,2 Joanne M. Morrisey,2 Benjamin A. Garcia,3 Akhil B. Vaidya,2 Joshua D. Rabinowitz,4 and Manuel Llinás1* 1Department of Molecular Biology & Lewis-Sigler Institute for Integrative Genomics, Princeton University, Princeton, NJ 08544 2Center for Molecular Parasitology, Drexel University College of Medicine, Philadelphia, PA 19129 3Department of Molecular Biology, Princeton University, Princeton, NJ 08544 4Department of Chemistry & Lewis-Sigler Institute for Integrative Genomics, Princeton University, Princeton, NJ 08544 Author Contributions K.O. cultured the parasites, collected and analyzed all LC-MS and GC-MS data; B.A.G. performed mass spectrometric analysis of histones. M.W.M. and J.M.M. carried out IDH localization studies. M.W.M. purified mitochondria and K.O. did biochemical assays. K.O., M.L., J.D.R., M.W.M., A.B.V. and B.A.G. designed the study; J.D.R. provided the metabolomic technology. M.L. and K.O. wrote the paper. All authors discussed the results and commented on the manuscript.